کریل (Euphausiids) [1] ( sg.: krill) سخت پوستان کوچک و منحصراً دریایی از راسته Euphausiacea هستند که در تمام اقیانوسهای جهان یافت میشوند. [2] نام "krill" از کلمه نروژی krill گرفته شده است ، به معنای "کوچک ماهی "، [3] که همچنین اغلب به گونههای ماهی نسبت داده میشود.
کریل به عنوان یک اتصال سطح تغذیه ای مهم در نظر گرفته می شود - نزدیک به انتهای زنجیره غذایی . آنها از فیتوپلانکتون و تا حدی زئوپلانکتون تغذیه می کنند و همچنین منبع اصلی غذا برای بسیاری از حیوانات بزرگتر هستند. در اقیانوس جنوبی ، یک گونه، کریل قطب جنوب ، زیست توده تخمینی حدود 379 میلیون تن را تشکیل می دهد ، [4] که آن را در میان گونه هایی با بیشترین زیست توده کل تشکیل می دهد. بیش از نیمی از این زیست توده هر ساله توسط نهنگ ها، فوک ها ، پنگوئن ها، پرندگان دریایی، ماهی مرکب و ماهی خورده می شود. اکثر گونههای کریل، مهاجرتهای عمودی روزانه زیادی را نشان میدهند ، بنابراین غذای شکارچیان در نزدیکی سطح در شب و در آبهای عمیقتر در طول روز فراهم میکنند.
کریل به صورت تجاری در اقیانوس جنوبی و در آب های اطراف ژاپن صید می شود. کل برداشت جهانی سالانه 150000 تا 200000 تن است که بیشتر آن از دریای اسکوشیا است . بیشتر صید کریل برای آبزی پروری و خوراک آکواریوم ، به عنوان طعمه در ماهیگیری ورزشی یا در صنعت داروسازی استفاده می شود. در ژاپن، فیلیپین و روسیه، کریل نیز برای مصرف انسان استفاده می شود و در ژاپن به اوکیامی (オキアミ) معروف است . آنها به عنوان کامارون در اسپانیا و فیلیپین خورده می شوند. در فیلیپین، کریل به نام آلامانگ نیز شناخته می شود و برای تهیه خمیر نمکی به نام باگونگ استفاده می شود .
راسته Euphausiacea شامل دو خانواده است . Euphausiidae فراوانتر شامل 10 جنس مختلف با مجموع 85 گونه است. از این میان، جنس Euphausia با 31 گونه، بزرگترین است. [5] خانواده کمتر شناخته شده، Bentheuphausiidae ، تنها یک گونه دارد ، Bentheuphausia amblyops ، یک کریل دریاچه ای که در آب های عمیق زیر 1000 متر (3300 فوت) زندگی می کند. این بدوی ترین گونه کریل موجود در نظر گرفته می شود. [6]
از سال 2013 [به روز رسانی]، راسته Euphausiacea به دلیل چندین ویژگی مورفولوژیکی حفاظت شده منحصر به فرد ( autapomorphy ) مانند آبشش های رشته ای برهنه و قفسه سینه های نازک [10] و مطالعات مولکولی، اعتقاد بر این است که تک فیلتیک است. [11] [12] [13]
نظریه های زیادی در مورد مکان راسته Euphausiacea وجود داشته است. از زمان اولین توصیف Thysanopode tricuspide توسط هانری میلن-ادواردز در سال 1830، شباهت قفسهپایان دوراموس آنها جانورشناسان را به گروهبندی euphausiids و Mysidacea در راسته Schizopoda که توسط یوهان اریک وستی بوآس در سال 188 به دو دسته جداگانه تقسیم شد، سوق داد. [14] بعدها، ویلیام توماس کالمن (1904) Mysidacea را در superorder Peracarida و euphausiids را در superorder Eucarida رتبهبندی کرد ، اگرچه حتی تا دهه 1930 از راسته Schizopoda حمایت میشد. [10] بعدها همچنین پیشنهاد شد که طبق رابرت گارنی و ایزابلا گوردون ، راسته Euphausiacea باید با Penaeidae (خانواده میگوها) در Decapoda بر اساس شباهت های رشدی گروه بندی شود . [15] [16] دلیل این بحث این است که کریلها برخی از ویژگیهای مورفولوژیکی دهپایان و برخی دیگر از میسیدها را به اشتراک میگذارند. [10]
مطالعات مولکولی آنها را بدون ابهام دسته بندی نکرده است، احتمالاً به دلیل کمبود گونه های کمیاب کلیدی مانند Bentheuphausia amblyops در کریل و Amphionides reynaudii در Eucarida. یک مطالعه از انحصار Eucarida (با Mysida پایه)، [17] گروه دیگر Euphausiacea با Mysida (Schizopoda)، [12] در حالی که گروه دیگری Euphausiacea را با Hoplocarida گروه می کند . [18]
جدول زمانی
هیچ فسیلی موجود را نمی توان به صراحت به Euphausiacea نسبت داد. برخی از گونه های منقرض شده eumalacostracan تصور می شود که Euphausiaceans مانند Anthracophausia ، Crangopsis - که اکنون به Aeschronectida (Hoplocarida) [8] اختصاص داده شده است - و Palaeomysis هستند . [19] تمام تاریخگذاری رویدادهای گونهزایی با روشهای ساعت مولکولی تخمین زده شد که آخرین جد مشترک خانواده کریل Euphausiidae (راست Euphausiacea منهای Bentheuphausia amblyops ) را در حدود 130 میلیون سال پیش در کرتاسه پایینتر میزیست . [12]
توزیع
کریل در سراسر جهان در تمام اقیانوس ها وجود دارد، اگرچه بسیاری از گونه های منفرد دارای توزیع بومی یا نرتیک ( به عنوان مثال، ساحلی) هستند. Bentheuphausia amblyops ، گونه ای دریاچه ای ، پراکندگی جهانی در زیستگاه اعماق دریا دارد . [20]
گونههای دارای پراکنش نریتیک شامل چهار گونه از جنس Nyctiphanes هستند . [22] آنها در امتداد مناطق بالارونده سیستم های فعلی کالیفرنیا ، هومبولت ، بنگوئلا و کاناریاس بسیار فراوان هستند . [23] [24] [25] گونه دیگری که فقط پراکنش نرتیک دارد E. crystallorophias است که در خط ساحلی قطب جنوب بومی است. [26]
گونه هایی با پراکنش بومی عبارتند از Nyctiphanes capensis ، که فقط در جریان بنگوئلا، [22] E. mucronata در جریان Humboldt، [27] و شش گونه Euphausia بومی اقیانوس جنوبی وجود دارد.
در قطب جنوب، هفت گونه شناخته شده است، [28] یکی در جنس Thysanoessa ( T. macrura ) و شش گونه در Euphausia . کریل قطب جنوب ( Euphausia superba ) معمولاً در اعماق 100 متر (330 فوت) زندگی می کند، [29] در حالی که کریل یخی ( Euphausia crystallorophias ) به عمق 4000 متر (13100 فوت) می رسد، اگرچه معمولاً در اعماق 6030-60 متر زندگی می کنند. متر (1000-2000 فوت). [30] کریل مهاجرتهای عمودی Diel (DVM) را در گروههای بزرگ انجام میدهد، و دادههای صوتی نشان دادهاند که این مهاجرتها تا عمق 400 متری میرود. [31] هر دو در عرض های جغرافیایی جنوب 55 درجه جنوبی ، با E. crystallorophias در جنوب 74 درجه جنوبی [32] و در مناطقی از یخ های بسته یافت می شوند . گونه های دیگر شناخته شده در اقیانوس جنوبی عبارتند از E. frigida ، E. longirostris ، E. triacantha و E. vallentini . [33]
آناتومی و مورفولوژی
آناتومی کریل با استفاده از Euphausia superba به عنوان مدل توضیح دادآبشش های کریل از بیرون قابل مشاهده هستند
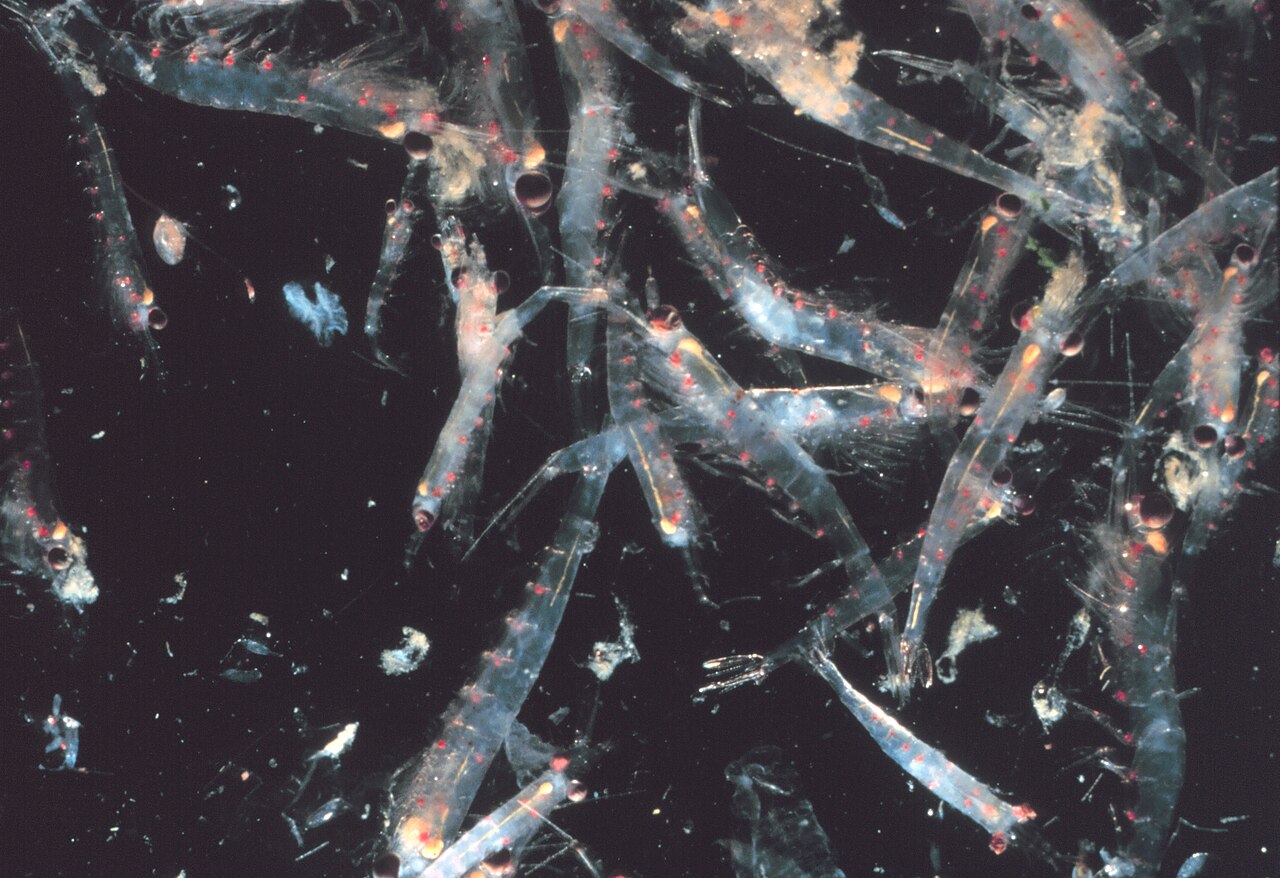
کریل ها سخت پوستان هستند و مانند همه سخت پوستان دارای اسکلت بیرونی کیتین هستند . آناتومی آنها شبیه به دهپایان استاندارد است و بدنشان از سه قسمت تشکیل شده است : سفالوتوراکس از سر و قفسه سینه که به هم جوش خوردهاند و شکم که ده زائده شنا را در خود دارد و فن دم تشکیل شده است . این پوسته بیرونی کریل در اکثر گونه ها شفاف است.
کریل دارای چشم های مرکب پیچیده است . برخی از گونه ها با استفاده از رنگدانه های غربالگری با شرایط مختلف نور سازگار می شوند . [34]
آنها دارای دو آنتن و چندین جفت پای قفسه سینه هستند که به آنها پریوپود یا قفسه سینه می گویند که به این دلیل به قفسه سینه چسبیده اند. تعداد آنها در جنس ها و گونه ها متفاوت است. این پاهای قفسه سینه شامل پاهای تغذیه و نظافت پاها است.
کریلها احتمالاً خواهر دهپایان هستند، زیرا همه گونهها دارای پنج جفت پا شنا به نام «شناگر» با پاهای دومی هستند که بسیار شبیه به پاهای خرچنگ یا خرچنگ آب شیرین است .
علیرغم داشتن ده شناگر که به عنوان پلوپود شناخته می شوند ، کریل را نمی توان ده پا دانست. آنها فاقد هر گونه پاهای واقعی مبتنی بر زمین هستند، زیرا تمام پاهای پریوپود آنها به پاهای نظافت و تغذیه کمکی تبدیل شده است. در Decapoda ، ده پریوپود فعال وجود دارد که نام آنها را می دهد. در حالی که در اینجا هیچ لوکوموتیو پریوپود باقی نمانده است . همچنین به طور مداوم ده پریوپود وجود ندارد .
بیشتر کریل ها در بزرگسالان حدود 1-2 سانتی متر (0.4-0.8 اینچ) طول دارند. تعداد کمی از گونه ها به اندازه های 6-15 سانتی متر (2.4-5.9 اینچ) رشد می کنند. بزرگترین گونه کریل، Thysanopoda spinicaudata ، در اعماق اقیانوس زندگی می کند . [35] کریل را می توان به راحتی از سایر سخت پوستان مانند میگوی واقعی با آبشش های قابل مشاهده بیرونی تشخیص داد . [36]
به جز Bentheuphausia amblyops ، کریل ها جانورانی بیولومنسانس هستند که اندام هایی به نام فوتوفور دارند که می توانند نور ساطع کنند. نور توسط یک واکنش نورتابی شیمیایی کاتالیز شده با آنزیم تولید می شود که در آن یک لوسیفرین (نوعی رنگدانه) توسط آنزیم لوسیفراز فعال می شود . مطالعات نشان میدهد که لوسیفرین بسیاری از گونههای کریل یک تتراپیرول فلورسنت است که مشابه لوسیفرین دینوفلاژلاتی است [37] و احتمالاً کریلها این ماده را خودشان تولید نمیکنند، بلکه آن را به عنوان بخشی از رژیم غذایی خود که حاوی دینوفلاژلات است، به دست میآورند. [38] فتوفورهای کریل اندام های پیچیده ای با لنزها و توانایی های تمرکز هستند و می توانند توسط عضلات چرخانده شوند. [39] عملکرد دقیق این اندام ها هنوز ناشناخته است. این امکانات عبارتند از جفت گیری، تعامل اجتماعی یا جهت گیری و به عنوان نوعی استتار ضد نور برای جبران سایه آنها در برابر نور بالای سر. [40] [41]
اکولوژی
فرآیندها در پمپ بیولوژیکی
فیتوپلانکتون CO 2 را که از اتمسفر به اقیانوس های سطحی حل شده است (90 Gt yr-1) به کربن آلی ذرات (POC) در طی تولید اولیه (~50 Gt C yr-1) تبدیل می کند. سپس فیتوپلانکتون ها توسط کریل ها و چرندگان زئوپلانکتون کوچک مصرف می شوند که به نوبه خود توسط سطوح تغذیه ای بالاتر طعمه می شوند. هر فیتوپلانکتون مصرف نشده، دانههایی را تشکیل میدهد، و همراه با گلولههای مدفوع زئوپلانکتون، به سرعت فرو میروند و از لایه مخلوط خارج میشوند (< 12 Gt C yr-1 14). کریل، زئوپلانکتون و میکروبها فیتوپلانکتونها را در سطح اقیانوس رهگیری میکنند و ذرات آواری را در عمق فرو میبرند، و این POC را به CO2 (کربن معدنی محلول، DIC) مصرف میکنند و تنفس میکنند ، به طوری که تنها بخش کوچکی از کربن تولید شده در سطح به اعماق اقیانوس فرو میرود. (یعنی اعماق بیش از 1000 متر). به عنوان تغذیه کریل و زئوپلانکتون کوچکتر، آنها همچنین ذرات را به صورت فیزیکی به قطعات کوچک، کندتر یا غیر غرق می کنند (از طریق تغذیه شلخته، کوپرورکسی در صورت تکه تکه شدن مدفوع)، و صادرات POC را به تاخیر می اندازند. این کربن آلی محلول (DOC) را مستقیماً از سلول ها یا به طور غیرمستقیم از طریق حل شدن باکتری (دایره زرد رنگ در اطراف DOC) آزاد می کند. سپس باکتری ها می توانند DOC را دوباره به DIC (CO 2 ، باغبانی میکروبی) معدنی کنند. کریل های مهاجر عمودی، زئوپلانکتون های کوچکتر و ماهی ها می توانند کربن را با مصرف POC در لایه سطحی در شب و متابولیزه کردن آن در اعماق اقامتگاه مزوپلژیک در طول روز به عمق انتقال دهند. بسته به تاریخچه حیات گونه، حمل و نقل فعال ممکن است به صورت فصلی نیز رخ دهد. اعداد داده شده شار کربن (Gt C yr-1) در جعبه های سفید و جرم کربن (Gt C) در جعبه های تیره هستند. [42]
تغذیه کردن
بسیاری از کریل ها فیدر هستند : [24] جلویی ترین زائده های آنها ، قفسه سینه ها، شانه های بسیار خوبی را تشکیل می دهند که با آن می توانند غذای خود را از آب فیلتر کنند. این فیلترها میتوانند در گونههایی (مانند Euphausia spp.) که عمدتاً از فیتوپلانکتونها ، بهویژه از دیاتومها ، که جلبکهای تک سلولی هستند، تغذیه میکنند، بسیار خوب باشند . کریل ها اکثراً همه چیزخوار هستند ، [43] اگرچه گونه های کمی گوشتخوار هستند و زئوپلانکتون های کوچک و لارو ماهی را شکار می کنند . [44]
کریل یک عنصر مهم از زنجیره غذایی آبزیان است . کریل ها تولید اولیه طعمه خود را به شکلی مناسب برای مصرف حیوانات بزرگتر تبدیل می کنند که نمی توانند مستقیماً از جلبک های کوچک تغذیه کنند. کریل شمالی و برخی گونههای دیگر دارای یک سبد فیلتر نسبتاً کوچک هستند و به طور فعال غلافها و زئوپلانکتونهای بزرگتر را شکار میکنند. [44]
شکار
بسیاری از حیوانات از کریل تغذیه می کنند، از حیوانات کوچکتر مانند ماهی یا پنگوئن گرفته تا حیوانات بزرگتر مانند فوک ها و نهنگ های بالین . [45]
اختلالات یک اکوسیستم که منجر به کاهش جمعیت کریل می شود می تواند اثرات گسترده ای داشته باشد. برای مثال، در طول شکوفایی کوکولیتوفور در دریای برینگ در سال 1998، [46] ، غلظت دیاتوم ها در ناحیه آسیب دیده کاهش یافت. کریل نمی تواند از کوکولیتوفورهای کوچکتر تغذیه کند، و در نتیجه جمعیت کریل (عمدتا E. pacifica ) در آن منطقه به شدت کاهش یافت. این به نوبه خود روی گونه های دیگر تأثیر گذاشت: جمعیت آب های برشی کاهش یافت. تصور می شد این حادثه یکی از دلایل عدم تخم ریزی ماهی قزل آلا در آن فصل بوده است . [47]
چندین مژک دار تک سلولی اندوپارازیتوئید از جنس Collinia می توانند گونه های کریل را آلوده کرده و جمعیت های آسیب دیده را ویران کنند. چنین بیماری هایی برای Thysanoessa inermis در دریای برینگ و همچنین برای E. pacifica ، Thysanoessa spinifera و T. gregaria در سواحل اقیانوس آرام آمریکای شمالی گزارش شده است . [48] [49] برخی از انگلهای خارجی از خانواده Dajidae ( ایزوپودهای epicaridean ) کریل (و همچنین میگو و mysids ) را مبتلا میکنند. یکی از این انگلها Oculophryxus bicaulis است که روی کریل Stylocheiron affine و S. longicorne یافت شد . خود را به ساقه چشم حیوان می چسباند و از سرش خون می مکد. ظاهراً از تولید مثل میزبان جلوگیری می کند، زیرا هیچ یک از حیوانات مبتلا به بلوغ نرسیده اند. [50]
تحقیقات اولیه نشان میدهد که کریل میتواند میکروپلاستیکهایی با قطر کمتر از 5 میلیمتر (0.20 اینچ) را هضم کند، آنها را شکسته و به شکل کوچکتر در محیط دفع کند. [52]
چرخه زندگی کریل علیرغم تغییرات جزئی در جزئیات از گونه ای به گونه دیگر، نسبتاً به خوبی درک شده است. [15] [24] پس از هچ کریل، آنها چندین مرحله لاروی را تجربه می کنند - ناپلیوس ، pseudometanauplius ، metanauplius ، calyptopsis ، و furcilia ، که هر کدام به مراحل فرعی تقسیم می شوند. مرحله pseudometanauplius انحصاری برای گونه هایی است که تخم های خود را در یک کیسه تخمگذار می گذارند: به اصطلاح "کیسه تخمگذار". لاروها در حین رشد به طور مکرر رشد می کنند و پوست اندازی می کنند و وقتی اسکلت بیرونی سفت و سخت خود را خیلی کوچک می کنند جایگزین می کنند. حیوانات کوچکتر بیشتر از حیوانات بزرگتر پوست اندازی می کنند. ذخایر زرده در بدن آنها لاروها را در مرحله متاناوپلیوس تغذیه می کند.
در مراحل کالیپتوپس، تمایز به اندازه کافی پیشرفت کرده است تا آنها بتوانند دهان و دستگاه گوارش ایجاد کنند و شروع به خوردن فیتوپلانکتون کنند. در آن زمان ذخایر زرده آنها تمام شده است و لاروها باید به منطقه فوتیک ، لایه های بالایی اقیانوس که جلبک ها در آن شکوفا می شوند، رسیده باشند. در طول مراحل furcilia، بخش هایی با جفت شناگر اضافه می شوند که از قسمت های جلویی شروع می شوند. هر جفت جدید فقط در مولت بعدی کاربردی می شود. تعداد قطعات اضافه شده در طول هر یک از مراحل furcilia ممکن است حتی در یک گونه بسته به شرایط محیطی متفاوت باشد. [53] پس از مرحله نهایی furcilia، یک نوجوان نابالغ به شکلی شبیه به بزرگسالان ظاهر می شود و متعاقباً غدد جنسی ایجاد می کند و از نظر جنسی بالغ می شود. [54]
تولید مثل
سر یک کریل ماده از گونه کیسه زایی Nematoscelis difficilis با کیسه مولدش. قطر تخم ها 0.3-0.4 میلی متر (0.012-0.016 اینچ) است.
در طول فصل جفت گیری، که بسته به گونه و آب و هوا متفاوت است، نر یک کیسه اسپرم را در دهانه تناسلی ماده (به نام thelycum ) رسوب می دهد. ماده ها می توانند چندین هزار تخم در تخمدان خود حمل کنند که ممکن است یک سوم توده بدن حیوان را تشکیل دهد. [55] کریل میتواند در یک فصل چندین مولد داشته باشد، با فواصل بین مولدین به ترتیب چند روز. [25] [56]
کریل از دو نوع مکانیسم تخم ریزی استفاده می کند. [25] 57 گونه از جنس Bentheuphausia ، Euphausia ، Meganyctiphanes ، Thysanoessa ، و Thysanopoda "پخش کننده های پخش" هستند: ماده تخم های بارور شده را در آب رها می کند، جایی که معمولاً غرق می شوند، پراکنده می شوند و به تنهایی هستند. این گونه ها عموماً در مرحله ناپلیوس 1 از تخم بیرون می آیند، اما اخیراً کشف شده است که گاهی اوقات به صورت متاناپلیوس یا حتی به صورت مراحل کالیپتوپیس بیرون می آیند. [57] 29 گونه باقیمانده از جنسهای دیگر "Sac Spawners" هستند، جایی که ماده تخمها را با خود حمل میکند و به عقبترین جفت قفسهپایان متصل میشود تا زمانی که به صورت متاناپلی از تخم بیرون بیایند، اگرچه برخی از گونهها مانند Nematoscelis difficilis ممکن است به صورت ناپلیوس یا ناپلیوس از تخم بیرون بیایند. pseudometanauplius. [58]
پوست اندازی
پوست اندازی زمانی اتفاق می افتد که یک نمونه از اسکلت بیرونی سفت و سخت خود رشد کند. حیوانات جوان که سریعتر رشد میکنند، بیشتر از حیوانات مسنتر و بزرگتر رشد میکنند. فرکانس پوست اندازی به طور گسترده ای برحسب گونه متفاوت است و حتی در یک گونه، تابع عوامل خارجی زیادی مانند عرض جغرافیایی، دمای آب و در دسترس بودن غذا است. به عنوان مثال ، گونه نیمه گرمسیری Nyctiphanes simplex ، دوره کلی بین دو تا هفت روزه دارد: لاروها به طور متوسط هر چهار روز یکبار پوست اندازی می کنند، در حالی که بچه ماهیان و بالغین به طور متوسط هر شش روز یکبار این کار را انجام می دهند. برای E. superba در دریای قطب جنوب، دوره های بین پوست اندازی بین 9 تا 28 روز بسته به دمای بین -1 تا 4 درجه سانتیگراد (30 و 39 درجه فارنهایت) مشاهده شده است، و برای Meganyctiphanes norvegica در دریای شمال دوره های بین پوست اندازی نیز از 9 و 28 روز اما در دمای بین 2.5 تا 15 درجه سانتیگراد (36.5 و 59.0 درجه فارنهایت) متغیر است. [59] E. superba میتواند اندازه بدن خود را زمانی که غذای کافی در دسترس نیست کاهش دهد، همچنین زمانی که اسکلت بیرونی آن بیش از حد بزرگ میشود، پوستاندازی میکند. [60] انقباض مشابهی برای E. pacifica نیز مشاهده شده است ، گونهای که در اقیانوس آرام از مناطق قطبی تا معتدل، به عنوان سازگاری با دمای غیرعادی بالا آب مشاهده شده است. انقباض برای سایر گونه های کریل منطقه معتدل نیز فرض شده است. [61]
طول عمر
برخی از گونه های کریل در عرض جغرافیایی بالا می توانند بیش از شش سال زندگی کنند (به عنوان مثال، Euphausia superba ). بقیه، مانند گونه های عرض جغرافیایی میانی Euphausia pacifica ، تنها دو سال زندگی می کنند. [7] طول عمر گونه های نیمه گرمسیری یا گرمسیری هنوز کوتاهتر است، به عنوان مثال، Nyctiphanes simplex ، که معمولاً تنها شش تا هشت ماه عمر می کند. [62]
ازدحام کردن
یک دسته کریل
اکثر کریل ها حیواناتی ازدحام هستند. اندازه و تراکم چنین دستههایی بسته به گونه و منطقه متفاوت است. برای Euphausia superba ، ازدحام به 10000 تا 60000 نفر در هر متر مکعب می رسد. [63] [64] ازدحام یک مکانیسم دفاعی است که شکارچیان کوچکتری را که میخواهند افراد را انتخاب کنند گیج میکند. در سال 2012، گاندومی و علوی الگوریتم تصادفی موفقی را برای مدلسازی رفتار ازدحام کریل ارائه کردند. این الگوریتم بر اساس سه عامل اصلی است: (1) حرکت ناشی از حضور افراد دیگر (2) فعالیت جستجوی غذا، و (iii) انتشار تصادفی." [65]
کریل معمولاً یک مهاجرت عمودی روزانه را دنبال می کند . فرض بر این است که آنها روز را در اعماق بیشتر می گذرانند و در طول شب به سمت سطح بالا می روند. هرچه عمیق تر می شوند، فعالیت خود را کاهش می دهند [66] ظاهراً برای کاهش برخورد با شکارچیان و صرفه جویی در انرژی. فعالیت شنا در کریل با پر بودن معده متفاوت است. حیوانات سیر شده ای که در سطح غذا تغذیه می کردند، کمتر شنا می کنند و بنابراین در زیر لایه مخلوط فرو می روند. [67] همانطور که آنها غرق می شوند، مدفوع تولید می کنند که نقشی در چرخه کربن قطب جنوب دارد . کریل با شکم خالی فعال تر شنا می کند و بنابراین به سمت سطح حرکت می کند.
مهاجرت عمودی ممکن است ۲ تا ۳ بار در روز اتفاق بیفتد. برخی از گونه ها (به عنوان مثال، Euphausia superba ، E. pacifica ، E. hanseni ، Pseudeuphausia latifrons ، و Thysanoessa spinifera ) در طول روز برای اهداف تغذیه و تولید مثل ازدحام سطحی تشکیل می دهند، حتی اگر چنین رفتاری خطرناک باشد زیرا آنها را به شدت در برابر شکارچیان آسیب پذیر می کند. [68]
مطالعات تجربی با استفاده از آرتمیا سالینا بهعنوان مدل نشان میدهد که مهاجرت عمودی کریل به طول چند صد متر، در گروههایی به عمق دهها متر، میتواند مجموعاً جتهای آب رو به پایین کافی ایجاد کند تا تأثیر قابلتوجهی بر اختلاط اقیانوس داشته باشد. [69]
ازدحام های متراکم می توانند باعث ایجاد جنون در تغذیه ماهی ها، پرندگان و شکارچیان پستانداران، به ویژه در نزدیکی سطح شوند. هنگامی که مختل می شود، یک دسته پراکنده می شود، و حتی برخی از افراد مشاهده شده اند که فورا پوست اندازی می کنند و اگزویا را به عنوان طعمه پشت سر می گذارند. [70]
کریلها معمولاً با سرعت 5 تا 10 سانتیمتر بر ثانیه (2 تا 3 طول بدن در ثانیه) شنا میکنند، [71] که از شناگرهای خود برای پیشرانش استفاده میکنند. مهاجرت های بزرگتر آنها در معرض جریان های اقیانوسی است. هنگامی که در معرض خطر قرار می گیرند، واکنش فراری به نام خرچنگ دریایی نشان می دهند - با تکان دادن ساختارهای دمی خود ، تلسون و اروپودها ، آنها نسبتاً سریع در آب به عقب حرکت می کنند و به سرعت هایی در محدوده 10 تا 27 طول بدن در ثانیه دست می یابند که برای کریل های بزرگ. مانند E. superba یعنی حدود 0.8 متر بر ثانیه (3 فوت بر ثانیه). [72] عملکرد شنای آنها بسیاری از محققین را بر آن داشته است تا کریل بالغ را به عنوان اشکال حیات میکرونکتونیک طبقه بندی کنند، یعنی حیوانات کوچکی که قادر به حرکت فردی در برابر جریانات (ضعیف) هستند. اشکال لارو کریل به طور کلی زئوپلانکتون در نظر گرفته می شود. [73]
چرخه های بیوژئوشیمیایی
نقش کریل قطب جنوب در چرخه های بیوژئوشیمیایی
کریل (به عنوان دسته ها و افراد) از فیتوپلانکتون در سطح (1) تغذیه می کند (1) تنها نسبتی را به عنوان دانه های گیاهی (2)، که به راحتی شکسته می شوند و ممکن است زیر ترموکلاین دائمی فرو نروند باقی می گذارد. کریل همچنین در حین تغذیه، گلوله های مدفوعی را آزاد می کند (3) که می تواند در اعماق دریا فرو برود اما می تواند مصرف شود (کوپروفاژی) و با فرود (4) توسط کریل، باکتری ها و زئوپلانکتون ها تخریب شود. در منطقه یخ حاشیه ای، شار گلوله های مدفوع می تواند به اعماق بیشتری برسد (5). کریل همچنین پوست اندازی را آزاد می کند که فرو می رود و به شار کربن کمک می کند (6). مواد مغذی در طی تغذیه نامرتب، دفع و هضم توسط کریل آزاد می شوند، مانند آهن و آمونیوم (7، برای سایر مواد مغذی آزاد شده به شکل 2 مراجعه کنید)، و اگر در نزدیکی سطح آزاد شوند، می توانند تولید فیتوپلانکتون و کاهش بیشتر CO2 اتمسفر را تحریک کنند . برخی از کریل های بالغ به طور دائم در اعماق ستون آب ساکن هستند و مواد آلی را در عمق مصرف می کنند (8). هر کربن (به عنوان ماده آلی یا CO 2 ) که در زیر ترموکلاین دائمی فرو میرود، از اختلاط فصلی خارج میشود و حداقل برای یک سال در اعماق اقیانوس ذخیره میشود (9). حرکات شنای کریل بالغ مهاجر که مهاجرت می کند می تواند آب غنی از مواد مغذی را از اعماق مخلوط کند (10) و تولید اولیه را بیشتر تحریک کند. سایر علوفه های کریل بالغ در کف دریا، CO2 تنفسی را در عمق آزاد می کنند و ممکن است توسط شکارچیان کفزی مصرف شوند (11). کریل لارو، که در اقیانوس جنوبی در زیر یخ دریا قرار دارد، تحت مهاجرت عمودی گسترده روزانه قرار می گیرد (12)، که به طور بالقوه CO2 را به زیر ترموکلاین دائمی منتقل می کند . کریل توسط بسیاری از شکارچیان از جمله نهنگ های بالین (13) مصرف می شود، که منجر به ذخیره بخشی از کربن کریل به عنوان زیست توده برای چندین دهه قبل از مرگ نهنگ، فرو رفتن در کف دریا و مصرف موجودات اعماق دریا می شود. [42]
هنگام پوست اندازی کریل، کلسیم، فلوراید و فسفر محلول را از اسکلت بیرونی آزاد می کنند (1). کیتین (مواد آلی) که اسکلت بیرونی را تشکیل می دهد به فرو رفتن شار ذرات آلی به اعماق اقیانوس کمک می کند. کریل بخشی از انرژی حاصل از مصرف فیتوپلانکتون یا سایر حیوانات را به عنوان دی اکسید کربن تنفس می کند (2)، هنگام شنا از آب های میانی/عمیق به سطح در گروه های بزرگ کریل مخلوط آب، که به طور بالقوه مواد مغذی را به آب های سطحی فقیر از مواد مغذی می آورد (3). آمونیوم و فسفات از آبشش ها و هنگام دفع همراه با کربن آلی محلول، نیتروژن (مثلا اوره) و فسفر (DOC، DON و DOP، 2 و 4) آزاد می شود. کریل گلولههای مدفوعی سریع غرقشونده حاوی کربن آلی، نیتروژن و فسفر (POC، PON و POP) و آهن را آزاد میکند که مورد دوم در هنگام شسته شدن به آبهای اطراف همراه با DOC، DON و DOP زیستی در دسترس است (5). [42]
استفاده های انسانی
بشقاب های انجماد عمیق از کریل قطب جنوب برای استفاده به عنوان خوراک حیوانات و مواد خام برای پخت و پز
تاریخچه برداشت
کریل حداقل از قرن نوزدهم به عنوان منبع غذایی برای انسان و حیوانات اهلی و احتمالاً قبل از آن در ژاپن، جایی که به اوکیامی معروف بود، برداشت میشد . ماهیگیری در مقیاس بزرگ در اواخر دهه 1960 و اوایل دهه 1970 توسعه یافت و اکنون فقط در آبهای قطب جنوب و در دریاهای اطراف ژاپن انجام می شود. از نظر تاریخی، بزرگترین کشورهای شیلات کریل ژاپن و اتحاد جماهیر شوروی، یا پس از انحلال شوروی، روسیه و اوکراین بودند . [77] برداشت به اوج خود رسید که در سال 1983 حدود 528000 تن تنها در اقیانوس جنوبی بود (که اتحاد جماهیر شوروی 93 درصد آن را بر عهده گرفت، اکنون به عنوان یک اقدام احتیاطی در برابر صید بی رویه مدیریت می شود. [78]
در سال 1993، دو رویداد باعث کاهش ماهیگیری کریل شد: روسیه از این صنعت خارج شد. و کنوانسیون حفاظت از منابع زنده دریایی قطب جنوب (CCAMLR) حداکثر سهمیه صید را برای بهره برداری پایدار از کریل قطب جنوب تعریف کرده است. پس از بررسی اکتبر 2011، کمیسیون تصمیم گرفت که این سهمیه را تغییر ندهد. [79]
صید سالانه قطب جنوب در حدود 100000 تن تثبیت شد که تقریباً یک پنجاهم سهمیه صید CCAMLR است. [80] عامل محدود کننده اصلی احتمالاً هزینه های بالا همراه با مسائل سیاسی و حقوقی بود. [81] شیلات ژاپنی در حدود 70000 تن اشباع شد. [82]
اگرچه کریل در سراسر جهان یافت می شود، ماهیگیری در اقیانوس های جنوبی ترجیح داده می شود زیرا کریل ها در این مناطق "قابل صید" و فراوان هستند. به ویژه در دریاهای قطب جنوب که بکر و بکر در نظر گرفته می شوند، آنها را "محصول پاک" می دانند. [77]
در سال 2018 اعلام شد که تقریباً همه شرکتهای ماهیگیری کریل که در قطب جنوب فعالیت میکنند، از سال 2020 فعالیتهای خود را در مناطق وسیعی در اطراف شبهجزیره قطب جنوب، از جمله «مناطق حائل» در اطراف مستعمرات پرورشدهنده پنگوئنها، رها خواهند کرد. [83]
اگرچه کل زیست توده کریل قطب جنوب ممکن است به وفور 400 میلیون تن باشد، تأثیر انسان بر این گونه اصلی در حال رشد است، با افزایش 39 درصدی در کل عملکرد ماهیگیری به 294000 تن طی سال های 2010-2014. [80] کشورهای عمده درگیر در برداشت کریل عبارتند از: نروژ (56٪ از کل صید در سال 2014)، جمهوری کره (19٪) و چین (18٪). [80]
کریل منبعی غنی از پروتئین و اسیدهای چرب امگا 3 است که در اوایل قرن بیست و یکم به عنوان غذای انسان، مکمل های غذایی به عنوان کپسول روغن، غذای دام و غذای حیوانات خانگی در دست توسعه هستند . [77] [79] [84] طعم کریل شور با طعم ماهی تا حدودی قوی تر از میگو است. برای مصرف انبوه و محصولاتی که به صورت تجاری تهیه می شوند، باید آنها را پوست کند تا اسکلت بیرونی غیر خوراکی از بین برود . [84]
کریل (و سایر میگوهای پلانکتونیک ، به ویژه Acetes spp.) بیشترین مصرف را در آسیای جنوب شرقی دارند، جایی که تخمیر می شوند (با پوسته سالم) و معمولاً برای تهیه خمیر میگو ریز آسیاب می شوند . میتوان آن را سرخکرده و همراه با برنج سفید میل کرد یا برای افزودن طعم اومامی به طیف گستردهای از غذاهای سنتی استفاده کرد. [86] [87] مایع حاصل از فرآیند تخمیر نیز به عنوان سس ماهی برداشت می شود . [88]
روباتیک با الهام از زیستی
کریل شناگران چابکی در رژیم اعداد رینولدز متوسط هستند ، که در آن راهحلهای زیادی برای روباتهای بدون خدمه زیر آب وجود ندارد و از پلتفرمهای رباتیک الهام گرفتهاند تا هم حرکت خود را مطالعه کنند و هم راهحلهای طراحی برای روباتهای زیر آب بیابند. [89]
↑ «Euphausiids (Krill)». دولت کانادا . شیلات و اقیانوس ها کانادا 6 آوریل 2022 . بازبینی شده در 18 آوریل 2024 . بسیاری از گونه های مختلف euphausiids در سواحل شرقی و غربی کانادا یافت می شوند.
↑ Crustacea: Euphausiacea - آکسفورد آکادمیک
↑ «کریل». دیکشنری ریشه شناسی آنلاین . بازبینی شده در 22 ژوئن 2010 .
^ A. Atkinson; V. Siegel; EA پاخوموف; MJ Jessopp; V. Loeb (2009). "ارزیابی مجدد کل زیست توده و تولید سالانه کریل قطب جنوب" (PDF) . بخش اول تحقیق در اعماق دریا . 56 (5): 727-740. Bibcode :2009DSRI...56..727A. doi :10.1016/j.dsr.2008.12.007.
↑ Siegel V (2011). Siegel V (ویرایش). "Euphausiidae Dana، 1852". پایگاه داده جهانی Euphausiacea . ثبت جهانی گونه های دریایی بازیابی شده در 25 نوامبر 2011 .
^ ab S. Nicol; Y. Endo (1999). "شیلات کریل: توسعه، مدیریت و پیامدهای اکوسیستم". منابع زندگی آبزی . 12 (2): 105-120. doi :10.1016/S0990-7440(99)80020-5. S2CID 84158071.
^ abcd آندریاس ماس; دیتر والوشک (2001). "توسعه لارو Euphausia superba Dana، 1852 و تجزیه و تحلیل فیلوژنتیکی Euphausiacea" (PDF) . هیدروبیولوژی . 448 : 143-169. doi :10.1023/A:1017549321961. S2CID 32997380. بایگانی شده از نسخه اصلی (PDF) در 18 جولای 2011.
↑ برنادت کازانووا (۱۹۸۴). "Phylogénie des Euphausiacés (Crustacés Eucarides)" [Phylogénie des Euphausiacés (Crustacea: Eucarida)]. Bulletin du Muséum National d'Histoire Naturelle (به فرانسوی). 4 : 1077-1089.
↑ abc Bernadette Casanova (2003). "Ordre des Euphausiacea Dana، 1852". سخت پوستان . 76 (9): 1083-1121. doi :10.1163/156854003322753439. JSTOR 20105650.
^ M. Eugenia D'Amato; گوردون دبلیو. هارکینز; تولیو د اولیویرا; پیتر R. Teske; مارک جی گیبونز (2008). "تاریخ مولکولی و جغرافیای زیستی کریل نرتیک Nyctiphanes" (PDF) . زیست شناسی دریایی . 155 (2): 243-247. doi :10.1007/s00227-008-1005-0. S2CID 17750015. بایگانی شده از نسخه اصلی (PDF) در 17 مارس 2012 . بازیابی شده در 4 جولای 2010 .
^ abc Simon N. Jarman (2001). "تاریخچه تکاملی کریل از تجزیه و تحلیل توالی rDNA زیرواحد بزرگ هسته ای استنباط شده است". مجله زیستی انجمن Linnean . 73 (2): 199-212. doi : 10.1111/j.1095-8312.2001.tb01357.x .
^ شین شن؛ هایکینگ وانگ؛ Minxiao Wang; بن لیو (2011). توالی کامل ژنوم میتوکندری Euphausia pacifica (Malacostraca: Euphausiacea) یک نظم ژنی جدید و تکرارهای پشت سر هم غیر معمول را نشان می دهد. ژنوم . 54 (11): 911-922. doi :10.1139/g11-053. PMID 22017501.
↑ یوهان اریک وستی بواس (۱۸۸۳). "Studien über die Verwandtschaftsbeziehungen der Malacostraken" [مطالعاتی در مورد روابط Malacostraca]. Morphologisches Jahrbuch (به آلمانی). 8 : 485-579.
↑ Trisha Spears، Ronald W. DeBry، Lawrence G. Abele & Katarzyna Chodyl (2005). بویکو، کریستوفر بی (ویرایش). "مونوفیلی پراکارید و فیلوژنی بین ردیفی استنتاج شده از توالی های DNA ریبوزومی زیرواحد کوچک هسته ای (Crustacea: Malacostraca: Peracarida)" (PDF) . مجموعه مقالات انجمن بیولوژیکی واشنگتن . 118 (1): 117-157. doi :10.2988/0006-324X(2005)118[117:PMAIPI]2.0.CO;2. S2CID 85557065.{{cite journal}}: CS1 maint: چندین نام: فهرست نویسندگان ( پیوند )
^ K. Meland; E. Willassen (2007). "تفرق "Mysidacea" (Crustacea)". فیلوژنتیک مولکولی و تکامل . 44 (3): 1083-1104. CiteSeerX 10.1.1.653.5935 . doi :10.1016/j.ympev.2007.02.009. PMID 17398121.
^ جی جی تورس؛ جی جی چیلدرس (1985). "تنفس و ترکیب شیمیایی euphausiid bathypelagic Bentheuphausia amblyops ". زیست شناسی دریایی . 87 (3): 267-272. doi :10.1007/BF00397804. S2CID 84486097.
↑ Volker Siegel (2011). "تیسانوئسا برانت، 1851". کرم ها . ثبت جهانی گونه های دریایی بازبینی شده در 18 ژوئن 2011 .
^ ab D'Amato، ME و همکاران. : "تاریخ مولکولی و جغرافیای زیستی کریل نرتیک Nyctiphanes بایگانی شده در 17 مارس 2012 در Wayback Machine "، در Marine Biology vol. 155، شماره 2 ، صفحات 243-247، اوت 2008.
↑ Volker Siegel (2011). V. Siegel (ویرایش). "Nyctiphanes Sars، 1883". پایگاه داده جهانی Euphausiacea . ثبت جهانی گونه های دریایی بازبینی شده در 18 ژوئن 2011 .
^ abc J. Mauchline; ال آر فیشر (1969). زیست شناسی اوفوسیدیان . پیشرفت در زیست شناسی دریایی جلد 7. مطبوعات دانشگاهی . شابک978-7-7708-3615-2.
^ abc Jaime Gómez-Gutiérrez; کارلوس جی رابینسون (2005). "زمان رشد جنینی، اولیه لاروی، مکانیسم جوجه کشی و دوره بین جوجه ریزی euphausiid Nyctiphanes simplex Hansen کیسه تخم ریزی". مجله تحقیقات پلانکتون . 27 (3): 279-295. doi : 10.1093/plankt/fbi003 .
^ SN Jarman; NG Elliott; S. Nicol; A. McMinn (2002). "تمایز ژنتیکی در کریل ساحلی قطب جنوب Euphausia crystallorophias". وراثت . 88 (4): 280-287. doi : 10.1038/sj.hdy.6800041 . PMID 11920136.
^ R. Escribano; وی. مارین; سی ایریبارن (2000). "توزیع Euphausia mucronata در منطقه بالا آمدن شبه جزیره Mejillones، شمال شیلی: تاثیر لایه حداقل اکسیژن". ساینتیا مارینا . 64 (1): 69-77. doi : 10.3989/scimar.2000.64n169 .
^ پی بروگمن. "Euphausia crystallorophias". راهنمای میدان زیر آب برای جزیره راس و صدای مک مردو، قطب جنوب . دانشگاه کالیفرنیا، سن دیگو .
↑ «Krill, Euphausia superba». MarineBio.org بازبینی شده در 25 فوریه 2009 .
↑ بیانکی، دانیله؛ Mislan، KAS (ژانويه 2016). "الگوهای جهانی زمان ها و سرعت های مهاجرت عمودی دیل از داده های صوتی". لیمنولوژی و اقیانوس شناسی . 61 (1). doi : 10.1002/lno.10219 .
^ A. Sala; م. عزلی; A. Russo (2002). "Krill of the Ross Sea: توزیع، فراوانی و جمعیت شناسی Euphausia superba و Euphausia crystallorophias در طول اکسپدیشن ایتالیایی قطب جنوب (ژانویه تا فوریه 2000)". ساینتیا مارینا . 66 (2): 123-133. doi : 10.3989/scimar.2002.66n2123 .
^ GW Hosie; M. Fukuchi; S. Kawaguchi (2003). "توسعه پیمایش ضبط کننده پلانکتون پیوسته اقیانوس جنوبی" (PDF) . پیشرفت در اقیانوس شناسی 58 (2-4): 263-283. Bibcode :2003PrOce..58..263H. doi :10.1016/j.pocean.2003.08.007.[ لینک مرده دائمی ]
^ ای. گتن. "Meganyctiphanes norvegica". دانشگاه لستر بایگانی شده از نسخه اصلی در 1 ژوئیه 2009 . بازبینی شده در 25 فوریه 2009 .
↑ ای. برینتون (1953). " Thysanopoda spinicauda ، سخت پوستان غول پیکر باتیپلاژیک euphausiid جدید، با نت های مقایسه ای در T. cornuta و T. egregia ". مجله آکادمی علوم واشنگتن . 43 : 408-412.
↑ «Euphausiacea». موسسه آبزی پروری و شیلات تاسمانی. بایگانی شده از نسخه اصلی در 30 سپتامبر 2009 . بازبینی شده در 6 ژوئن 2010 .
↑ O. Shimomura (1995). "نقش دو جزء بسیار ناپایدار F و P که در بیولومینسانس میگوهای euphausiid دخیل هستند". مجله بیولومینسانس و نورتابی شیمیایی . 10 (2): 91-101. doi :10.1002/bio.1170100205. PMID 7676855.
^ جی سی دانلپ؛ JW Hastings; O. Shimomura (1980). "واکنش متقابل بین سیستم های ساطع کننده نور موجودات دوردست: نوع جدیدی از ترکیب ساطع کننده نور". مجموعه مقالات آکادمی ملی علوم . 77 (3): 1394–1397. Bibcode :1980PNAS...77.1394D. doi : 10.1073/pnas.77.3.1394 . JSTOR 8463. PMC 348501 . PMID 16592787.
^ پی جی شاه ماهی؛ EA Widder (2001). "بیولومینسانس در پلانکتون و نکتون". در JH Steele; SA Thorpe; KK Turekian (ویرایشگران). دایره المعارف علوم اقیانوسی . جلد 1. انتشارات آکادمیک ، سن دیگو. صص 308-317. شابک978-0-12-227430-5.
^ اس ام لیندسی; MI Latz (1999). شواهد تجربی برای سایه زنی درخشنده توسط برخی سخت پوستان euphausiid . نشست علوم آبزیان انجمن لیمنولوژی و اقیانوس شناسی آمریکا (ASLO). سانتافه
↑ سونکه جانسن (2005). "قرمز و سیاه: بیولومینسانس و رنگ حیوانات در اعماق دریا" (PDF) . زیست شناسی تلفیقی و تطبیقی . 4 (2): 234-246. doi : 10.1093/icb/45.2.234 . PMID 21676767. S2CID 247718. بایگانی شده از نسخه اصلی (PDF) در 2 اکتبر 2005.
^ abcd Cavan، EL، Belcher، A.، Atkinson، A.، Hill، SL، Kawaguchi، S.، McCormack، S.، Meyer، B.، Nicol، S.، Ratnarajah، L.، Schmidt، K. و Steinberg، DK (2019) "اهمیت کریل قطب جنوب در چرخه های بیوژئوشیمیایی". ارتباطات طبیعت ، 10 (1): 1-13. doi :10.1038/s41467-019-12668-7.مطالب از این منبع کپی شده است، که تحت مجوز Creative Commons Attribution 4.0 International در دسترس است.
^ جی سی کریپس؛ A. Atkinson (2000). "ترکیب اسیدهای چرب به عنوان شاخص گوشتخواری در کریل قطب جنوب، Euphausia superba ". مجله کانادایی شیلات و علوم آبزیان . 57 (S3): 31-37. doi :10.1139/f00-167.
↑ ام جی شرام (10 اکتبر 2007). «کریل کوچک: غولها در زنجیره غذایی دریایی». برنامه ملی پناهگاه دریایی NOAA . بازبینی شده در 4 ژوئن 2010 .
^ جی وایر (1999). "جریانهای در حال تغییر دریای برینگ را سایه جدیدی از آبی رنگ می کند." رصدخانه زمین NOAA . بازیابی شده در 15 ژوئن 2005 .
^ RD Brodeur; GH Kruse; PA Livingston; جی. والترز; J. Ianelli; GL Swartzman; M. Stepanenko; T. Wyllie-Echeverria (1998). پیش نویس گزارش کارگاه بین المللی FOCI در مورد شرایط اخیر در دریای برینگ . NOAA ص 22-26.
↑ جی روچ (17 ژوئیه 2003). "دانشمندان قاتل کریل مرموز را کشف کردند". اخبار نشنال جئوگرافیک بایگانی شده از نسخه اصلی در 24 ژوئیه 2003.
^ جی. گومز-گوتیرز; WT Peterson; A. de Robertis; RD Brodeur (2003). "مرگ و میر انبوه کریل ناشی از مژه های انگل". علم . 301 (5631): 339. doi :10.1126/science.1085164. PMID 12869754. S2CID 28471713.
^ جی دی شیلدز. جی. گومز-گوتیرز (1996). " Oculophryxus bicaulis ، یک جنس و گونه جدید انگلی ایزوپود داجید بر روی euphausiid Stylocheiron affine Hansen". مجله بین المللی انگل شناسی . 26 (3): 261-268. doi :10.1016/0020-7519(95)00126-3. PMID 8786215.
↑ Rusty Dornin (6 ژوئیه 1997). "جمعیت کریل قطب جنوب در حال کاهش است". سی ان ان بازبینی شده در 18 ژوئن 2011 .
داوسون ، آماندا ال. کاواگوچی، پس؛ کینگ، کاترین ک. تاونسند، کتی آ. کینگ، رابرت؛ هیوستون، ویلهلمینا ام. بنگتسون نش، سوزان ام (2018). "تبدیل میکروپلاستیک به نانوپلاستیک از طریق قطعه قطعه شدن گوارشی توسط کریل قطب جنوب". ارتباطات طبیعت . 9 (1): 1001. Bibcode :2018NatCo...9.1001D. doi :10.1038/s41467-018-03465-9. PMC 5843626 . PMID 29520086.
↑ ام دی نایت (1984). "تغییر در مورفوژنز لارو در جمعیت خلیج کالیفرنیای جنوبی Euphausia pacifica از زمستان تا تابستان، 1977-1978" (PDF) . گزارش CalCOFI XXV بایگانی شده از نسخه اصلی (PDF) در 3 اوت 2019 . بازبینی شده در 5 نوامبر 2017 .
^ آر ام راس; ال بی کوتین (1986). "کریل قطب جنوب چقدر مولد هستند؟" علوم زیستی . 36 (4): 264-269. doi :10.2307/1310217. JSTOR 1310217.
↑ Janine Cuzin-Roudy (2000). "تولید فصلی، تخم ریزی متعدد، و باروری در کریل شمالی، Meganyctiphanes norvegica ، و کریل قطب جنوب، Euphausia superba ". مجله کانادایی شیلات و علوم آبزیان . 57 (S3): 6-15. doi : 10.1139/f00-165.
↑ جی. گومز-گوتیرز (2002). "مکانیسم جوجه ریزی و هچ با تاخیر تخم های سه گونه euphausiid در حال تخم ریزی در شرایط آزمایشگاهی". مجله تحقیقات پلانکتون . 24 (12): 1265-1276. doi : 10.1093/plankt/24.12.1265 .
^ ای. برینتون; MD Ohman; AW Townsend; ام دی نایت; AL Bridgeman (2000). Euphausiids of the World Ocean. مجموعه CD-ROM پایگاه داده جهانی تنوع زیستی، Springer Verlag . شابک978-3-540-14673-5. بایگانی شده از نسخه اصلی در 26 فوریه 2012 . بازبینی شده در 4 دسامبر 2009 .
↑ F. Buchholz (2003). "آزمایشهایی بر روی فیزیولوژی کریل جنوبی و شمالی، Euphausia superba و Meganyctiphanes norvegica ، با تاکید بر پوستریزی و رشد - مروری". رفتار و فیزیولوژی دریایی و آب شیرین . 36 (4): 229-247. Bibcode :2003MFBP...36..229B. doi :10.1080/10236240310001623376. S2CID 85121989.
^ H.-C. ساق پا S. Nicol (2002). "استفاده از رابطه بین قطر چشم و طول بدن برای تشخیص اثرات گرسنگی طولانی مدت بر کریل Euphausia superba قطب جنوب". سری پیشرفت اکولوژی دریایی 239 : 157-167. Bibcode :2002MEPS..239..157S. doi : 10.3354/meps239157 .
^ بی. مارینوویچ; M. Mangel (1999). "کریل می تواند به عنوان یک سازگاری اکولوژیکی با محیط های موقتی نامطلوب کوچک شود" (PDF) . نامه های بوم شناسی . 2 : 338-343.
↑ جی جی گومز (۱۹۹۵). "الگوهای توزیع، فراوانی و پویایی جمعیت euphausiidsNyctiphanes simplex و Euphausia eximia در سواحل غربی باخا کالیفرنیا، مکزیک" (PDF) . سری پیشرفت اکولوژی دریایی 119 : 63-76. Bibcode :1995MEPS..119...63G. doi : 10.3354/meps119063 .
^ یو کیلز; پی مارشال (1995). "Der Krill, wie er schwimmt und frisst – neue Einsichten mit neuen Methoden ("کریل قطب جنوب - نحوه شنا و تغذیه - بینش های جدید با روش های جدید ")". در I. Hempel; G. Hempel (ویرایشات). Biologie der Polarmeere – Erlebnisse und Ergebnisse ( زیست شناسی تجربیات و نتایج اقیانوس های قطبی ) . فیشر ورلاگ ص 201-210. شابک978-3-334-60950-7.
^ R. Piper (2007). حیوانات خارق العاده: دایره المعارف حیوانات کنجکاو و غیرمعمول . پرس گرین وود شابک978-0-313-33922-6.
↑ گندمی، ق. علوی، ق (1391). "کریل هرد: یک الگوریتم جدید بهینه سازی الهام گرفته از زیست". ارتباطات در علوم غیرخطی و شبیه سازی عددی . 17 (12): 4831-4845. Bibcode :2012CNSNS..17.4831G. doi :10.1016/j.cnsns.2012.05.010.
^ JS Jaffe; MD Ohmann; A. de Robertis (1999). "برآورد سونار سطوح فعالیت روزانه Euphausia pacifica در ورودی سانیچ" (PDF) . مجله کانادایی شیلات و علوم آبزیان . 56 (11): 2000–2010. doi :10.1139/cjfas-56-11-2000. S2CID 228567512. بایگانی شده از نسخه اصلی (PDF) در 20 ژوئیه 2011.
↑ Geraint A. Tarling; مگنوس ال جانسون (2006). "سیری به کریل احساس غرق شدن می دهد". زیست شناسی فعلی . 16 (3): 83-84. doi : 10.1016/j.cub.2006.01.044 . PMID 16461267.
↑ دن هاوارد (2001). "کریل" (PDF) . در Herman A. Karl; جان ال چین; ادوارد اوبر؛ پیتر اچ استافر; جیمز دبلیو هندلی دوم (ویرایشها). فراتر از دروازه طلایی - اقیانوس شناسی، زمین شناسی، زیست شناسی، و مسائل زیست محیطی در خلیج فارالون. سازمان زمین شناسی ایالات متحده صص 133-140. بخشنامه 1198 . بازیابی شده در 8 اکتبر 2011 .
^ دی. هوارد. "کریل در پناهگاه دریایی ملی بانک کوردل". اداره ملی اقیانوسی و جوی بازیابی شده در 15 ژوئن 2005 .
^ دیوید A. Demer; استفان جی کونتی (2005). مدل جدید مقاومت هدف نشان دهنده کریل بیشتر در اقیانوس جنوبی است. مجله علوم دریایی ICES . 62 (1): 25-32. doi : 10.1016/j.icesjms.2004.07.027 .
↑ یو کیلز (1982). "رفتار شنا، عملکرد شنا و تعادل انرژی کریل قطب جنوب Euphausia superba". BIOMASS Scientific Series 3, BiomaSS Research Series : 1-122. بایگانی شده از نسخه اصلی در 2 ژوئن 2020 . بازبینی شده در 11 نوامبر 2017 .
^ S. Nicol; Y. Endo (1997). "شیلات کریل جهان". مقاله فنی شیلات فائو 367 .
↑ Ratnarajah, L., Bowie, AR, Lannuzel, D., Meiners, KM and Nicol, S. (2014) "نقش بیوژئوشیمیایی نهنگ های بالین و کریل در چرخه مواد مغذی اقیانوس جنوبی". PLOS ONE , 9 (12): e114067. doi :10.1371/journal.pone.0114067
^ Hopkins, TL, Ainley, DG, Torres, JJ, Lancraft, TM, 1993. ساختار استوایی در آبهای آزاد منطقه یخ حاشیه ای در منطقه تلاقی اسکوشیا ودل در طول بهار (1983). زیست شناسی قطبی 13، 389-397.
↑ Lancraft, TM, Relsenbichler, KR, Robinson, BH, Hopkins, TL, Torres, JJ, 2004. یک جامعه میکرونکتونی و ماکروزوپلانکتون تحت سلطه کریل در کروکر پاساژ، قطب جنوب با تخمین شکار ماهی. تحقیقات دریای عمیق II 51، 2247-2260.
↑ abc Grossman، Elizabeth (14 ژوئیه 2015). دانشمندان بررسی می کنند که آیا کریل باید از شکار بیش از حد انسان محافظت شود یا خیر. رادیو عمومی بین المللی (PRI) . بازبینی شده در 1 آوریل 2017 .
↑ «شیلات کریل و پایداری: کریل قطب جنوب (Euphausia superba)». کمیسیون حفاظت از منابع زیست دریایی قطب جنوب. 23 آوریل 2015 . بازبینی شده در 1 آوریل 2017 .
^ ab Schiermeier، Q (2010). بوم شناسان از بحران کریل قطب جنوب می ترسند. طبیعت . 467 (7311): 15. doi : 10.1038/467015a . PMID 20811427.
^ abc "Krill - زیست شناسی، بوم شناسی و ماهیگیری". کمیسیون حفاظت از منابع زیست دریایی قطب جنوب. 28 آوریل 2015 . بازبینی شده در 1 آوریل 2017 .
↑ مینتورن جی رایت (1987). "مالکیت قطب جنوب، منابع زنده و معدنی آن". مجله حقوق و محیط زیست . 4 (2): 49-78.
^ S. Nicol; جی فاستر (2003). "روندهای اخیر در ماهیگیری کریل قطب جنوب". منابع زندگی آبزی . 16 : 42-45. doi :10.1016/S0990-7440(03)00004-4.
↑ جاش، گاباتیس (10 ژوئیه 2018). صنعت ماهیگیری کریل از پناهگاه عظیم اقیانوس قطب جنوب برای محافظت از پنگوئن ها، فوک ها و نهنگ ها حمایت می کند. مستقل . بازیابی شده در 10 جولای 2018 .
^ ab "چرا کریل؟". مرکز علوم شیلات جنوب غربی، اداره ملی اقیانوسی و جوی ایالات متحده. 22 نوامبر 2016 . بازبینی شده در 1 آوریل 2017 .
↑ Cheeseman MA (22 ژوئیه 2011). "روغن کریل: نامه پاسخ آژانس GRAS اطلاعیه شماره GRN 000371". FDA آمریکا . بازبینی شده در 3 ژوئن 2015 .
↑ عموری، م. (1978). "شیلات زئوپلانکتون جهان: بررسی". زیست شناسی دریایی . 48 (3): 199-205. doi :10.1007/BF00397145. S2CID 86540101.
^ پونگستکول، جاکسوما؛ بنژاکول، سوتاوات؛ سامپاواپل، پونانی؛ اوساکو، کازوفومی؛ Faithong، Nandhsha (17 سپتامبر 2014). "ترکیب شیمیایی و خواص فیزیکی خمیر میگو شور (کاپی) تولید تایلند". تحقیقات بین المللی آبزیان . 6 (3): 155-166. doi : 10.1007/s40071-014-0076-4 .
^ آبه، کنجی؛ سوزوکی، کنجی؛ هاشیموتو، کانهیسا (1979). "استفاده از کریل به عنوان ماده سس ماهی". نیپون سویسان گاکایشی . 45 (8): 1013-1017. doi : 10.2331/suisan.45.1013 .
↑ اولیویرا سانتوس، سارا؛ تاک، نیلز; سو، یونکسینگ؛ کوئنکا-خیمنز، فرانسیسکو؛ مورالس لوپز، اسکار؛ گومز والدز، پی آنتونیو; ام ویلهلموس، مونیکا (13 ژوئن 2023). "Pleobot: یک راه حل رباتیک مدولار برای شنای متاکرونال". گزارش های علمی 13 (1): 9574. Bibcode :2023NatSR..13.9574O. doi :10.1038/s41598-023-36185-2. PMC 10264458 . PMID 37311777.
برینتون، ادوارد : "Euphausiids از آبهای جنوب شرقی آسیا". گزارش ناگا جلد 4، قسمت 5. لا جولا: دانشگاه کالیفرنیا، موسسه اقیانوس شناسی اسکریپس، 1975.
کانوی، DVP؛ سفید، RG; هیوگ-دیت-سیلز، جی. Galienne، CP; Robins, DB: راهنمای زئوپلانکتونهای ساحلی و سطحی جنوب غربی اقیانوس هند بایگانیشده در 23 اکتبر 2012 در Wayback Machine ، Order Euphausiacea، انتشار گاه به گاه انجمن بیولوژیکی دریایی بریتانیا، شماره 15، پلیموث، انگلستان، 220 .
اورسون، آی (ویرایش): کریل: زیستشناسی، اکولوژی و شیلات . آکسفورد، علم بلک ول؛ 2000. ISBN 0-632-05565-0 .
Hamner, William M. (مه 1984). "کریل - فضل دست نخورده از دریا؟". نشنال جئوگرافیک . جلد 165، شماره 5. صص 626-642. ISSN 0027-9358. OCLC 643483454.
Mauchline, J.: Euphausiacea: Adults بایگانی شده در 15 مه 2011 در Wayback Machine , Conseil International pour l'Exploration de la Mer, 1971. برگه های شناسایی برای کریل بزرگسالان با بسیاری از نقشه های خطی. فایل پی دی اف 2 مگابایت
Mauchline, J.: Euphausiacea: Larvae بایگانی شده در 19 آوریل 2012 در Wayback Machine , Conseil International pour l'Exploration de la Mer, 1971. برگه های شناسایی مراحل لاروی کریل با بسیاری از نقشه های خطی. فایل پی دی اف 3 مگابایت
تت، پ.: زیست شناسی Euphausiids ، یادداشت های سخنرانی از یک دوره در سال 2003 در زیست شناسی دریایی از دانشگاه ناپیر .
تت، پ.: بیولومینسانس ، یادداشت های سخنرانی از نسخه 1999/2000 همان دوره.
لینک های خارجی
در ویکیانبار پروندههایی مربوط به کریل موجود است.










{kind=link}