_(10144905255).jpg/1280px-Anthomyiidae_(female)_(10144905255).jpg)
مگس ها حشراتی از راسته دوپترا هستند که نام آن از واژه یونانی δι- دی- «دو» و πτερόν pteron «بال» گرفته شده است. حشرات از این راسته فقط از یک جفت بال برای پرواز استفاده می کنند، بال های عقبی به اندام های حسی مکانیکی پیشرفته معروف به هالتر تبدیل شده اند که به عنوان حسگرهای سرعت بالا در حرکت چرخشی عمل می کنند و به دوپتران ها اجازه می دهند هوازی پیشرفته انجام دهند. Diptera یک راسته بزرگ است که شامل حدود 1,000,000 گونه از جمله مگس اسب ، مگس جرثقیل ، مگس شناور ، پشه و غیره تخمین زده شده است، اگرچه تنها حدود 125000 گونه توصیف شده است .
مگس ها دارای یک سر متحرک، با یک جفت چشم مرکب بزرگ ، و قطعات دهانی هستند که برای سوراخ کردن و مکیدن (پشه، مگس سیاه و مگس دزد)، یا برای لپ زدن و مکیدن در گروه های دیگر طراحی شده اند. چیدمان بالها به آنها قدرت مانور زیادی در پرواز میدهد و پنجهها و بالشتکهای روی پاها آنها را قادر میسازد تا به سطوح صاف بچسبند. مگس ها دچار دگردیسی کامل می شوند . تخمها اغلب روی منبع غذایی لارو گذاشته میشوند و لاروها که فاقد اندام واقعی هستند، در یک محیط محافظتشده و اغلب در داخل منبع غذایی خود رشد میکنند. گونههای دیگر تخمزا هستند و لاروهای هچ شده یا جوجهآور را بهجای تخمها روی لاشه ، سرگین، مواد در حال پوسیدگی یا زخمهای باز پستانداران قرار میدهند. شفیره یک کپسول سخت است که وقتی بالغ برای انجام آن آماده باشد از آن خارج می شود. مگس ها در بزرگسالی عمر کوتاهی دارند.
دوپترا یکی از راسته های اصلی حشرات است که از اهمیت اکولوژیکی و انسانی قابل توجهی برخوردار است. مگس ها گرده افشان های مهمی هستند و بعد از زنبورها و بستگان Hymenopteran آنها در رتبه دوم قرار دارند . مگس ها ممکن است یکی از اولین گرده افشان های تکاملی باشند که مسئول گرده افشانی اولیه گیاهان بوده اند . مگسهای میوه به عنوان ارگانیسمهای مدل در تحقیقات استفاده میشوند ، اما پشهها ناقل مالاریا ، تب دنگی ، تب نیل غربی ، تب زرد ، آنسفالیت و سایر بیماریهای عفونی هستند . و مگسهای خانگی ، مشابه انسانها در سراسر جهان، بیماریهای ناشی از غذا را منتشر میکنند . مگسها میتوانند آزاردهنده باشند، بهویژه در برخی از نقاط جهان که میتوانند به تعداد زیاد ایجاد شوند، وزوز کنند و روی پوست یا چشمها بنشینند و گاز بگیرند یا به دنبال مایعات بگردند. مگسهای بزرگتر مانند مگس تسهسه و کرمهای حلزونی آسیب اقتصادی قابل توجهی به گاو وارد میکنند. لارو مگس بادکن که به نام جنتلز شناخته می شود و سایر لاروهای دوپتران که به طور کلی با نام ماگت شناخته می شوند، به عنوان طعمه ماهیگیری ، به عنوان غذای حیوانات گوشتخوار و در پزشکی در دبریدمان برای تمیز کردن زخم ها استفاده می شود .
دیپتران ها هولومتابولان ها هستند ، حشراتی که دچار دگردیسی ریشه ای می شوند. آنها در کنار Mecoptera ، Siphonaptera ، Lepidoptera و Trichoptera به Mecopterida تعلق دارند . [3] [4] داشتن یک جفت بال، بیشتر مگس های واقعی را از سایر حشرات با نام "مگس" متمایز می کند. با این حال، برخی از مگس های واقعی مانند Hippoboscidae (مگس شپش) به طور ثانویه بدون بال شده اند. [5] [6]
کلادوگرام نمایانگر دیدگاه اجماع فعلی است . [7]
.jpg/1280px-Fossil_insect_Diptera,_Brachycera_in_Baltic_amber._Age_50_Mill._years_(the_Lower_Eocene).jpg)
اولین دیپتران های واقعی شناخته شده مربوط به تریاس میانی (حدود 240 میلیون سال پیش) هستند و در طول تریاس میانی و پسین گسترش یافتند . [8] گیاهان گلدار مدرن تا دوره کرتاسه (حدود 140 میلیون سال پیش) ظاهر نشدند ، بنابراین دیپتران های اصلی باید منبع تغذیه متفاوتی به غیر از شهد داشته باشند . بر اساس جذابیت بسیاری از گروه های مگس مدرن به قطرات براق، پیشنهاد شده است که آنها ممکن است از عسلک تولید شده توسط حشرات مکنده شیره که در آن زمان فراوان بودند تغذیه می کردند، و قطعات دهانی دیپتران به خوبی برای نرم کردن و چسباندن مگس ها سازگار هستند. باقی مانده های پوسته دار [9] کلادهای پایه در دوپترا شامل Deuterophlebiidae و Nymphomyiidae معمایی هستند . [10] تصور می شود که سه قسمت از تشعشعات تکاملی بر اساس سابقه فسیلی رخ داده است. بسیاری از گونه های جدید دوپتراهای پایین تر در تریاس ، حدود 220 میلیون سال پیش، توسعه یافتند. بسیاری از Brachycera پایین در ژوراسیک ، حدود 180 میلیون سال پیش ظاهر شدند. تابش سوم در میان شیزوفورا در آغاز پالئوژن ، 66 میلیون سال پیش رخ داد. [10]
موقعیت فیلوژنتیکی دوپترا بحث برانگیز بوده است. انحصار حشرات هولومتابولوس از دیرباز پذیرفته شده است و راستههای اصلی آنها به صورت Lepidoptera، Coleoptera، Hymenoptera و Diptera ایجاد شده است و روابط بین این گروهها است که مشکلاتی را ایجاد کرده است. به طور گسترده تصور می شود که دوپترا به همراه Lepidoptera (پروانه ها و پروانه ها)، Trichoptera (کادیس مگس)، Siphonaptera (کک)، Mecoptera (مگس عقرب) و احتمالاً strepsiptera (مگس های بال های پیچ خورده) عضوی از Mecopterida هستند . Diptera با Siphonaptera و Mecoptera در Antliophora گروه بندی شده است، اما این توسط مطالعات مولکولی تایید نشده است. [11]

دوپتراها به طور سنتی به دو زیرمجموعه تقسیم می شدند، Nematocera و Brachycera که با تفاوت در آنتن ها متمایز می شدند. نماتوسراها با بدن دراز و شاخک های پر از چند قطعه و اغلب پر مانند پشه ها و مگس های جرثقیل شناسایی می شوند. Brachycera بدن گردتر و آنتن های بسیار کوتاه تری دارد. [12] [13] مطالعات بعدی نماتوسرا را به عنوان غیر منوفیلتیک با فیلوژنی های مدرن شناسایی کردند که براکیسرا را در گروه هایی از گروه هایی که قبلا در نماتوسرا قرار می گرفتند قرار می داد. ساخت یک درخت فیلوژنتیک موضوع تحقیقات مداوم بوده است. کلادوگرام زیر بر اساس پروژه FLYTREE است. [14] [15]

مگس ها اغلب فراوان هستند و تقریباً در تمام زیستگاه های زمینی جهان به غیر از قطب جنوب یافت می شوند. آنها شامل بسیاری از حشرات آشنا مانند مگس خانگی، مگس بادکن، پشه، پشه، مگس سیاه، میخ و مگس میوه هستند. بیش از 150000 مورد به طور رسمی توصیف شده است و تنوع گونه های واقعی بسیار بیشتر است و مگس ها از بسیاری از نقاط جهان هنوز به طور فشرده مورد مطالعه قرار نگرفته اند. [16] [17] زیررده Nematocera شامل حشرات به طور کلی کوچک و باریک با آنتن های بلند مانند پشه ها، پشه ها، مگس ها و مگس های جرثقیل است، در حالی که Brachycera شامل مگس های گسترده تر و قوی تر با آنتن های کوتاه است. بسیاری از لاروهای نماتوسران آبزی هستند. [18] تخمین زده میشود که در مجموع حدود 19000 گونه دوپترا در اروپا، 22000 گونه در ناحیه نزدیکی، 20000 گونه در منطقه افروتروپیکال، 23000 گونه در منطقه شرقی و 19000 گونه در منطقه استرالیا وجود دارد. [19] در حالی که بیشتر گونه ها پراکنش محدودی دارند، تعداد کمی مانند مگس خانگی ( Musca domestica ) جهان وطنی هستند. [20] Gauromydas heros ( Asiloidea )، با طول تا 7 سانتی متر (2.8 اینچ)، به طور کلی به عنوان بزرگترین مگس در جهان در نظر گرفته می شود، [21] در حالی که کوچکترین آنها Euryplatea nanaknihali است که با 0.4 میلی متر (0.016) در) کوچکتر از یک دانه نمک است. [22]
Brachycera از نظر اکولوژیکی بسیار متنوع است، بسیاری از آنها در مرحله لاروی شکارچی و برخی انگلی هستند. حیوانات انگلی شامل نرم تنان ، شپش های چوبی ، هزارپاها ، حشرات، پستانداران ، [19] و دوزیستان هستند . [23] مگس ها بعد از Hymenoptera (زنبورها، زنبورها و خویشاوندان) دومین گروه بزرگ گرده افشان ها هستند. در محیط های مرطوب و سردتر، مگس ها به عنوان گرده افشان اهمیت قابل توجهی دارند. در مقایسه با زنبورها، آنها به غذای کمتری نیاز دارند زیرا نیازی به تهیه بچه های خود ندارند. بسیاری از گلهایی که شهد کم دارند و آنهایی که گرده افشانی تله دارند به مگس بستگی دارند. [24] تصور می شود که برخی از اولین گرده افشان های گیاهان ممکن است مگس ها بوده باشند. [25]
بیشترین تنوع حشرات کیسه صفرا در میان مگس ها، عمدتاً در خانواده Cecidomyiidae (موج های صفراوی) یافت می شود. [26] بسیاری از مگس ها (مهمتر از خانواده Agromyzidae) تخم های خود را در بافت مزوفیل برگ ها می گذارند و لاروها بین سطوح تغذیه می کنند و تاول ها و مین ها را تشکیل می دهند. [27] برخی از خانوادهها قارچخوار یا قارچخوار هستند. اینها شامل غارهای Mycetophilidae (پنه های قارچی) می شود که لاروهای آن تنها دوپترا با نورتابی زیستی هستند. Sciaridae همچنین تغذیه کننده قارچ هستند. برخی از گیاهان توسط مگس های تغذیه کننده قارچ که از گل های نر آلوده به قارچ بازدید می کنند گرده افشانی می شوند. [28]
لاروهای Megaselia scalaris (Phoridae) تقریباً همه چیزخوار هستند و موادی مانند رنگ و جلا دهنده کفش را مصرف می کنند. [29] مگس Exorista mella (واکر) ژنرالیست و پارازیتوئید میزبان های مختلف در نظر گرفته می شود. [30] لارو مگسهای ساحلی (Ephydridae) و برخی Chironomidae در محیطهای شدید از جمله یخچالهای طبیعی ( Diamesa sp.، Chironomidae [31] )، چشمههای آب گرم، آبفشانها، حوضچههای شور، حوضچههای گوگرد، مخازن سپتیک و حتی نفت خام زنده میمانند. Helaeomyia Petrolei [31] ). [19] مگسهای شناور بالغ (Syrphidae) به دلیل تقلیدشان به خوبی شناخته شدهاند و لاروها شیوههای زندگی متنوعی را اتخاذ میکنند که از جمله آنها میتوان به لاشخورهای غیرطبیعی در داخل لانه حشرات اجتماعی اشاره کرد. [32] برخی از براکیسران ها آفات کشاورزی هستند، برخی حیوانات و انسان ها را گاز می گیرند و خون آنها را می مکند و برخی ناقل بیماری ها هستند. [19]
مگس ها برای حرکت هوایی مناسب هستند و معمولاً بدنی کوتاه و ساده دارند. اولین تگمای مگس، سر، چشمها، شاخکها و قسمتهای دهان را نشان میدهد (لابروم، لابیوم، فک پایین و ماگزیلا قسمتهای دهان را تشکیل میدهند). تگما دوم، قفسه سینه ، دارای بال است و شامل ماهیچه های پرواز در بخش دوم است که به شدت بزرگ شده است. بخش های اول و سوم به ساختارهای یقه مانند تبدیل شده اند و بخش سوم دارای هالترها هستند که به تعادل حشره در طول پرواز کمک می کنند. سومین تاگما شکم است که از 11 بخش تشکیل شده است که برخی از آنها ممکن است به هم جوش بخورند و 3 قسمت عقبی آن برای تولید مثل اصلاح شده است. [33] [34] برخی از دیپتران ها تقلید هستند و تنها با بازرسی بسیار دقیق می توان آنها را از مدل های آنها متمایز کرد. نمونه ای از آن Spilomyia longicornis است که مگس است اما شبیه زنبور vespid است .
.jpg/1280px-Tabanus_atratus,_U,_Face,_MD_2013-08-21-16.06.31_ZS_PMax_(9599360121).jpg)
_(25234095874).jpg/1280px-Tephrochlamys_rufiventris_(female_head)_(25234095874).jpg)
مگس ها دارای یک سر متحرک با یک جفت چشم مرکب بزرگ در طرفین سر و در بیشتر گونه ها، سه اوسل کوچک در بالای سر دارند. چشمهای مرکب ممکن است نزدیک به هم یا کاملاً از هم جدا باشند و در برخی موارد به یک ناحیه پشتی و یک ناحیه شکمی تقسیم میشوند، شاید برای کمک به رفتار انبوهی. آنتن ها به خوبی توسعه یافته اند اما متغیر هستند و در خانواده های مختلف مانند نخ، پر یا شانه مانند هستند. قطعات دهانی مانند مگس سیاه، پشه و مگس سارق برای سوراخ کردن و مکیدن و مانند بسیاری از گروههای دیگر برای لپ زدن و مکیدن مناسب هستند. [34] مگس اسب ماده از فک پایین و فک بالا مانند چاقو برای ایجاد یک برش صلیبی شکل در پوست میزبان استفاده میکند و سپس خونی را که جریان مییابد در دست میگیرد. روده شامل دیورتیکول های بزرگی است که به حشره اجازه می دهد تا مقدار کمی مایع را بعد از غذا ذخیره کند. [35]
برای کنترل مسیر بصری، میدان جریان نوری مگس ها توسط مجموعه ای از نورون های حساس به حرکت تجزیه و تحلیل می شود. [36] تصور میشود که زیرمجموعهای از این نورونها در استفاده از جریان نوری برای تخمین پارامترهای حرکت خود، مانند انحراف، چرخش، و ترجمه جانبی نقش دارند. [37] تصور میشود که سایر نورونها در تحلیل محتوای خود صحنه بصری، مانند جدا کردن چهرهها از زمین با استفاده از اختلاف منظر حرکتی، نقش دارند. [38] [39] نورون H1 مسئول تشخیص حرکت افقی در سراسر میدان بینایی مگس است و به مگس اجازه میدهد تا اصلاحات حرکتی تثبیتکننده را در اواسط پرواز با توجه به انحراف ایجاد کند و هدایت کند. [40] Ocelli در تشخیص تغییرات در شدت نور نگران است و مگس را قادر می سازد تا به سرعت به نزدیک شدن به یک جسم واکنش نشان دهد. [41]
مانند سایر حشرات، مگس ها دارای گیرنده های شیمیایی هستند که بو و طعم را تشخیص می دهند و گیرنده های مکانیکی که به لمس پاسخ می دهند. بخش سوم شاخک ها و کف فک بالا حامل گیرنده های بویایی اصلی هستند، در حالی که گیرنده های چشایی در لابیوم، حلق، پاها، حاشیه بال ها و اندام تناسلی زن قرار دارند، [42] که مگس ها را قادر می سازد با راه رفتن روی آن غذای خود را بچشند. گیرنده های چشایی در زنان در نوک شکم اطلاعاتی را در مورد مناسب بودن مکان برای تخمگذاری دریافت می کنند. [41] مگس هایی که از خون تغذیه می کنند ساختارهای حسی خاصی دارند که می توانند تشعشعات مادون قرمز را تشخیص دهند و از آنها برای خانه در میزبان خود استفاده کنند . [43] برخی از مگسهای تاچینید (Ormiinae) که پارازیتوئیدهای جیرجیرکهای بوتهای هستند ، گیرندههای صوتی دارند تا به آنها کمک کند تا میزبانهای آوازخوان خود را پیدا کنند. [44]

دوپتراها یک جفت بال جلویی روی مزوتوراکس و یک جفت هالتر یا بالهای عقبی کوچک شده روی متاتوراکس دارند . سازگاری بیشتر برای پرواز، کاهش تعداد عقدههای عصبی ، و تمرکز بافت عصبی در قفسه سینه است، ویژگی که در زیر نظم بسیار مشتق شده Muscomorpha شدیدترین است. [35] برخی از مگسها مانند Nycteribiidae و Streblidae انگلهای نابجا به دلیل از دست دادن بالهای خود و عدم پرواز استثنایی هستند. تنها دسته دیگر از حشرات که دارای یک جفت بال واقعی و کاربردی هستند، علاوه بر هر شکلی از هالترها، Strepsiptera هستند . برخلاف مگسها، استرپسیپتراها بالهای خود را روی مزوتوراکس و بالهای پرواز خود را روی متاتوراکس نگه میدارند. [45] هر یک از شش پای مگس دارای ساختار حشرهای معمولی از کوکسا، تروکانتر، استخوان ران، تیبیا و تارسوس است که در بیشتر موارد تارسوس به پنج تارسومر تقسیم میشود . [34] در نوک اندام یک جفت پنجه وجود دارد، و بین آنها ساختارهای بالشتکی به نام پولویلی وجود دارد که چسبندگی را ایجاد می کند. [46]
شکم تنوع قابل توجهی را در بین اعضای راسته نشان می دهد. این شامل یازده بخش در گروه های ابتدایی و ده بخش در گروه های مشتق شده تر است که بخش دهم و یازدهم با هم ترکیب شده اند. [47] دو یا سه بخش آخر برای بازتولید اقتباس شده است. هر بخش از یک اسکلریت پشتی و شکمی تشکیل شده است که توسط یک غشای الاستیک به هم متصل شده اند. در برخی از زنان، اسکلریت ها به شکل یک تخمگذار تلسکوپی منعطف می چرخند . [34]

مگس ها به دلیل وجود هالترها توانایی مانور زیادی در طول پرواز دارند. اینها به عنوان اندام های ژیروسکوپی عمل می کنند و به سرعت در زمان با بال ها نوسان می کنند. آنها با ارائه بازخورد سریع به ماهیچه های فرمان بال به عنوان یک سیستم تعادل و هدایت عمل می کنند و مگس هایی که از هولتر خود محروم هستند قادر به پرواز نیستند. بال ها و هالترها همزمان حرکت می کنند اما دامنه ضربان هر بال مستقل است و به مگس اجازه می دهد به طرفین بچرخد. [48] بالهای مگس به دو نوع ماهیچه متصل میشوند، ماهیچههایی که برای نیرو دادن به آن استفاده میشوند و دستهای دیگر برای کنترل خوب. [49]
مگس ها تمایل دارند در یک خط مستقیم پرواز کنند، سپس قبل از ادامه مسیر مستقیم، تغییر جهت سریع ایجاد می کنند. تغییرات جهت را ساکاد می نامند و معمولاً شامل زاویه 90 درجه است که در 50 میلی ثانیه به دست می آید. آنها توسط محرک های بصری هنگامی که مگس یک شی را مشاهده می کند آغاز می شود، سپس اعصاب عضلات فرمان را در قفسه سینه فعال می کنند که باعث تغییر کوچکی در حرکت بال می شود که گشتاور کافی برای چرخش ایجاد می کند. با تشخیص این موضوع در طی چهار یا پنج ضربه بال، هالترها یک چرخش معکوس ایجاد می کنند و مگس به سمتی جدید حرکت می کند. [50]
مگس ها دارای رفلکس های سریعی هستند که به فرار آنها از شکارچیان کمک می کند، اما سرعت پرواز پایدار آنها کم است. مگس های Dolichopodid از جنس Condylostylus در کمتر از 5 میلی ثانیه به فلاش دوربین با پرواز پاسخ می دهند. [ 51] در گذشته، بر اساس برآوردی که چارلز تاونسند در سال 1927 به صورت بصری انجام داد ، مگس ربات آهو، Cepenemyia ، یکی از سریعترین حشرات است . در هر ساعت، مرتباً تکرار شد تا زمانی که ایروینگ لانگمویر نشان داد که از نظر فیزیکی غیرممکن و همچنین نادرست است. لانگمویر سرعت تخمینی 25 مایل در ساعت را پیشنهاد کرد. [53] [54] [55]
اگرچه بیشتر مگس ها در نزدیکی زمین زندگی می کنند و پرواز می کنند، تعداد کمی از آنها در ارتفاعات پرواز می کنند و تعداد کمی مانند Oscinella (Chloropidae) شناخته شده است که توسط باد در ارتفاعات تا 2000 فوت و در فواصل طولانی پراکنده می شوند. [56] برخی از مگسهای شناور مانند Metasyrphus corollae شناخته شدهاند که در پاسخ به جهش جمعیت شتهها، پروازهای طولانی انجام میدهند. [57]
نرهای گونههای مگس مانند Cuterebra ، بسیاری از مگسهای شناور، [58] مگسهای زنبور عسل (Bombyliidae) [59] و مگسهای میوه (Tephritidae) [60] سرزمینهایی را حفظ میکنند که در آنها در تعقیب هوایی برای بیرون راندن نرهای مزاحم و سایر گونهها شرکت میکنند. [61] در حالی که این مناطق ممکن است توسط نرهای منفرد نگهداری شوند، برخی از گونهها، مانند A. freeborni ، [62] با تعداد زیادی از نرها که در نمایشها جمع میشوند، لکها را تشکیل میدهند . [60] برخی از مگسها فضای هوایی را حفظ میکنند و برخی دیگر ازدحامهای متراکم را تشکیل میدهند که مکان ثابتی را با توجه به نشانهها حفظ میکنند. بسیاری از مگس ها در هنگام پرواز جفت گیری می کنند. [63]
.jpg/1280px-Anthomyiidae_sp._1_(aka).jpg)
دوپتراها با چهار مرحله زندگی متمایز - تخم، لارو، شفیره و بالغ، یک دگردیسی کامل را پشت سر می گذارند.
در بسیاری از مگس ها، مرحله لاروی طولانی است و بزرگسالان ممکن است عمر کوتاهی داشته باشند. بیشتر لاروهای دیپتران در محیط های محافظت شده رشد می کنند. بسیاری از آنها آبزی هستند و برخی دیگر در مکان های مرطوب مانند مردار، میوه، مواد سبزیجات، قارچ ها و در مورد گونه های انگلی در داخل میزبان خود یافت می شوند. آنها معمولاً کوتیکول های نازکی دارند و در صورت قرار گرفتن در معرض هوا خشک می شوند. به غیر از Brachycera ، اکثر لاروهای دیپتران دارای کپسولهای سر اسکلرو شده هستند که ممکن است به قلابهای دهان باقیمانده تبدیل شوند. با این حال، Brachycera دارای کپسولهای سر نرم و ژلاتینهای است که اسکلریتها در آنها کاهش یافته یا از بین رفتهاند. بسیاری از این لاروها سر خود را به داخل قفسه سینه خود می برند. [34] [64] مارپیچ ها در لارو و شفیره هیچ وسیله مکانیکی بسته داخلی ندارند. [65]

برخی دیگر از تمایزات تشریحی بین لاروهای نماتوسرا و براکیسرا وجود دارد . به خصوص در Brachycera، مرز کمی بین قفسه سینه و شکم دیده می شود، اگرچه این مرز ممکن است در بسیاری از نماتوسراها، مانند پشه ها، قابل مشاهده باشد. در Brachycera، سر لارو به وضوح از بقیه بدن قابل تشخیص نیست و تعداد کمی اسکلریت وجود دارد. به طور غیررسمی، به این گونه لاروهای براکیسران، ماگوت می گویند، [66] اما این اصطلاح فنی نیست و اغلب به طور کلی برای لارو مگس یا لارو حشرات بی تفاوت به کار می رود. چشم ها و شاخک های لارو براکیسران کاهش یافته یا وجود ندارد و شکم نیز فاقد زائده هایی مانند سرسی است . این فقدان ویژگی، سازگاری با مواد غذایی مانند مردار، ریزه های پوسیده یا بافت میزبان اطراف انگل های داخلی است . [35] لاروهای نماتوسران عموماً چشمها و شاخکهای خوبی دارند، در حالی که لاروهای براکیسران کاهش یافته یا اصلاح شدهاند. [67]
لاروهای دیپتران هیچ گونه مفصلی و "پای واقعی" ندارند، [64] اما برخی از لاروهای دوپتران، مانند گونه های Simuliidae ، Tabanidae و Vermileonidae ، دارای پرولگ هایی هستند که برای نگه داشتن روی یک بستر در آب جاری، بافت میزبان یا طعمه سازگار هستند. [68] اکثر دوپترانها تخمزا هستند و دستههایی از تخمگذاری میکنند، اما برخی از گونهها تخمزا هستند ، جایی که لاروها قبل از جوجهشدن یا زندهزا شدن در داخل تخمها شروع به رشد میکنند، لاروها در بدن مادر بیرون میآیند و بالغ میشوند. . اینها به ویژه در گروه هایی یافت می شوند که لاروهای وابسته به منابع غذایی دارند که عمر کوتاهی دارند یا برای دوره های کوتاهی در دسترس هستند. [69] این در برخی از خانواده ها مانند Sarcophagidae گسترده است. در Hylemya strigosa (Anthomyiidae) لارو قبل از جوجه ریزی تا سن دوم پوست اندازی می کند و در Termitoxenia (Phoridae) ماده ها دارای کیسه های جوجه کشی هستند و یک لارو سن سوم کاملاً رشد یافته توسط بالغ رسوب می کند و تقریباً بلافاصله بدون مرحله لارو تغذیه آزادانه شفیره می شود. . مگس تسه تسه (و همچنین دیگر Glossinidae، Hippoboscidae، Nycteribidae و Streblidae) صفات زندهداری آدنوتروفیک را نشان میدهد . یک تخمک بارور شده در مجرای تخمک باقی می ماند و لارو در حال رشد از ترشحات غدد تغذیه می کند. هنگامی که به طور کامل رشد کرد، ماده یک نقطه با خاک نرم پیدا می کند و لارو راه خود را از مجرای تخمک خارج می کند، خود را دفن می کند و شفیره می شود. برخی از مگسها مانند Lundstroemia parthenogenetica (Chironomidae) با پارتنوژنز thelytokous تولید مثل میکنند و برخی از مگسهای صفراوی دارای لاروهایی هستند که میتوانند تخم تولید کنند ( paedogenesis ). [70] [71]
شفیره ها اشکال مختلفی دارند. در برخی از گروه ها، به ویژه نماتوسرا، شفیره حد واسط بین لارو و بالغ است. این شفیره ها به عنوان "ابکتکت" توصیف می شوند، که زائده های آینده را به عنوان ساختارهایی که به بدن شفیرگ می چسبند قابل مشاهده هستند. سطح بیرونی شفیره ممکن است خارهای چرمی و خرس، ویژگی های تنفسی یا پاروهای حرکتی باشد. در گروه های دیگر که به عنوان "کوآرکتات" توصیف می شوند، زائده ها قابل مشاهده نیستند. در اینها، سطح بیرونی یک شفیره است که از آخرین پوست لارو تشکیل شده است و شفیره واقعی درون آن پنهان است. هنگامی که حشره بالغ آماده بیرون آمدن از این کپسول سخت و مقاوم در برابر خشک شدن است، ساختاری شبیه بادکنک را روی سر خود باد می کند و مجبور به خروج از آن می شود. [34]
مرحله بزرگسالی معمولا کوتاه است، عملکرد آن فقط جفت گیری و تخم گذاری است. اندام تناسلی مگسهای نر به درجات متفاوتی از موقعیتی که در سایر حشرات یافت میشود میچرخند. [72] در برخی مگسها، این چرخش موقت در طول جفتگیری است، اما در برخی دیگر، پیچش دائمی اندامها است که در مرحله شفیرگی رخ میدهد. این پیچ خوردگی ممکن است منجر به قرار گرفتن مقعد در زیر اندام تناسلی شود، یا در مورد پیچ خوردگی 360 درجه، مجرای اسپرم به دور روده پیچیده شود و اندام های خارجی در موقعیت معمول خود قرار گیرند. هنگامی که مگس ها جفت گیری می کنند، مگس نر در ابتدا روی ماده به سمت همان جهت پرواز می کند، اما سپس در جهت مخالف می چرخد. این امر نر را مجبور می کند به پشت بخوابد تا اندام تناسلی او با دستگاه تناسلی ماده درگیر بماند یا پیچ خوردگی اندام تناسلی نر به نر اجازه می دهد در حالی که ایستاده است جفت گیری کند. این منجر به این می شود که مگس ها نسبت به اکثر حشرات توانایی تولید مثل بیشتری داشته باشند و بسیار سریعتر. مگس ها به دلیل توانایی آنها در جفت گیری موثر و سریع در طول فصل جفت گیری در جمعیت های بزرگی وجود دارند. [35] گروههای ابتداییتر در طول ازدحام در هوا جفت میگیرند، اما بیشتر گونههای پیشرفتهتر با پیچش 360 درجه روی یک بستر جفت میشوند. [73]

به عنوان حشرات همه جا حاضر، دیپتران ها نقش مهمی در سطوح مختلف تغذیه ای هم به عنوان مصرف کننده و هم به عنوان طعمه بازی می کنند. در برخی از گروه ها لاروها بدون تغذیه کامل رشد می کنند و در برخی دیگر بالغ ها تغذیه نمی کنند. لاروها می توانند گیاهخوار، لاشخور، تجزیه کننده، شکارچی یا انگل باشند که مصرف مواد آلی در حال پوسیدگی یکی از رایج ترین رفتارهای تغذیه ای است. میوه یا ریزهها همراه با میکروارگانیسمهای مرتبط مصرف میشوند، یک فیلتر غربال مانند در حلق برای متمرکز کردن ذرات استفاده میشود، در حالی که لاروهای گوشتخوار دارای قلابهایی در دهان هستند که به خرد کردن غذا کمک میکنند. لارو برخی از گروه ها از بافت های زنده گیاهان و قارچ ها تغذیه می کنند و برخی از آنها آفات جدی محصولات کشاورزی هستند. برخی از لاروهای آبزی لایههای جلبکهایی را که در زیر آب روی سنگها و گیاهان تشکیل میشوند مصرف میکنند. بسیاری از لاروهای پارازیتوئید در داخل رشد می کنند و در نهایت بندپایان دیگر را می کشند، در حالی که لاروهای انگلی ممکن است به میزبان مهره داران حمله کنند. [34]
در حالی که بسیاری از لاروهای دیپتران آبزی هستند یا در مکان های خشکی محصور زندگی می کنند، اکثر افراد بالغ در بالای زمین زندگی می کنند و قادر به پرواز هستند. آنها عمدتاً از شهد یا ترشحات گیاهی یا حیوانی مانند عسلک تغذیه میکنند که قسمتهای دهانی آنها برای آنها مناسب است. برخی از مگس ها دارای فک پایین عملکردی هستند که ممکن است برای گاز گرفتن استفاده شود. مگسهایی که از خون مهرهداران تغذیه میکنند، سبکهای تیزی دارند که پوست را سوراخ میکنند، با برخی از گونهها بزاق ضد انعقاد دارند که قبل از جذب خونی که در جریان است، برگشت داده میشود. در این فرآیند بیماری های خاصی قابل انتقال هستند. مگس ربات (Oestridae) برای انگلی کردن پستانداران تکامل یافته است. بسیاری از گونه ها چرخه زندگی خود را در داخل بدن میزبان خود کامل می کنند. [74] لارو چند گروه مگس (Agromyzidae، Anthomyiidae، Cecidomyiidae) قادر به القای گال گیاهان هستند. برخی از لاروهای دیپتران برگ معدنی هستند. لاروهای بسیاری از خانواده های براکیسران شکاری هستند. در بسیاری از گروههای دیپتران، ازدحام کردن یکی از ویژگیهای زندگی بزرگسالان است که ابرهای حشرات در مکانهای خاصی جمع میشوند. این حشرات اکثراً نر هستند و ازدحام ممکن است به این منظور باشد که مکان آنها برای ماده ها قابل مشاهده باشد. [34]
قسمتهای دهانی اکثر دوقچههای بالغ اصلاح شده تا مایع اسفنجی شود. بزرگسالان بسیاری از گونههای مگس (مانند Anthomyia sp.، Steganopsis melanogaster ) که از غذای مایع تغذیه میکنند، مایع را با رفتاری به نام «حباب زدن» پس میگیرند که تصور میشود به حشرات کمک میکند تا آب را تبخیر کنند و غذا را غلیظ کنند [75] یا احتمالاً تا با تبخیر خنک شود. [76] برخی از دوقلوهای بالغ به دلیل kleptoparasitism مانند اعضای Sarcophagidae شناخته شده اند. میلیتوگرامها به دلیل عادتشان به دنبال کردن زنبورها و دزدیدن طعمههای نیشخورده یا تخمگذاری درون آنها به «مگسهای ماهوارهای» معروف هستند. فوریدها، میلیشیدها و جنس بنگالیا به دزدیدن غذای حمل شده توسط مورچه ها معروف هستند. [77] بزرگسالان افیدرا هیانس در زیر آب علوفه می خرند و موهای آبگریز خاصی دارند که حباب هوا را به دام می اندازد که به آنها اجازه می دهد زیر آب نفس بکشند. [78]

مگس ها در تمام مراحل رشد توسط حیوانات دیگر خورده می شوند. تخمها و لاروها توسط حشرات دیگر انگلی میشوند و توسط بسیاری از موجودات خورده میشوند، برخی از آنها در تغذیه از مگسها تخصص دارند اما بیشتر آنها آنها را به عنوان بخشی از یک رژیم غذایی مخلوط مصرف میکنند. پرندگان، خفاش ها، قورباغه ها، مارمولک ها، سنجاقک ها و عنکبوت ها از جمله شکارچیان مگس ها هستند. [79] بسیاری از مگس ها شباهت های تقلیدی ایجاد کرده اند که به محافظت از آنها کمک می کند. تقلید باتسی با بسیاری از مگسهای شناور شبیه زنبورها و زنبورها، [80] [81] مورچهها [82] و برخی از گونههای مگس میوه تفریتی شبیه به عنکبوت گسترده است. [83] برخی از گونههای مگسهای هواگر میرمکوفیل هستند - جوانهای آنها در لانههای مورچهها زندگی و رشد میکنند. آنها با تقلید بوهای شیمیایی که توسط اعضای کلنی مورچه ها ایجاد می شود از مورچه ها محافظت می شوند. [84] مگسهای زنبور Bombyliid مانند Bombylius major بدن کوتاه، گرد، پشمالو و مشخصاً شبیه زنبور عسل هستند زیرا برای شهد از گلها بازدید میکنند و احتمالاً شبیه زنبورهای باتسی نیز هستند. [85]
در مقابل، مگس سرکه ، گونهای از مگسها در جنس مگس سرکه ، فاقد دستهای از هموسیتها است که در سایر گونههای مورد مطالعه مگس سرکه وجود دارد ، که منجر به ناتوانی در دفاع در برابر حملات انگلی، نوعی نقص ایمنی ذاتی میشود. [86]

مگس ها نقش های نمادین مختلفی را در فرهنگ های مختلف بازی می کنند. اینها شامل نقش های مثبت و منفی در دین می شود. در آیین سنتی ناواهو ، مگس بزرگ یک موجود روحی مهم است. [۸۷] [۸۸] [۸۹] در شیطانشناسی مسیحی ، بلزبوب یک مگس شیطانی، «ارباب مگسها» و خدای فلسطینیان است . [90] [91] [92]
مگس ها از سومر باستان در ادبیات ظاهر شده اند . [93] در یک شعر سومری، مگس به الهه Inanna کمک می کند ، زمانی که شوهرش Dumuzid توسط شیاطین گالا تعقیب می شود . [93] در نسخه های بین النهرینی از افسانه سیل ، اجساد مرده شناور بر روی آب ها با مگس ها مقایسه می شوند. [93] بعدها، گفته می شود که خدایان "مانند مگس" در اطراف هدیه قهرمان اوتناپیشتیم ازدحام می کنند. [93] مگس ها بر روی مهرهای بابلی قدیم به عنوان نماد نرگال ، خدای مرگ ظاهر می شوند. [93] مهرههای لاجورد به شکل مگس اغلب در بین النهرین باستان همراه با انواع دیگر جواهرات مگس پوشیده میشدند. [93]
در مصر باستان، مگس ها در طلسم ها ظاهر می شوند و به عنوان یک جایزه نظامی برای شجاعت و سرسختی ظاهر می شوند، به این دلیل که آنها همیشه در هنگام ضربه زدن برمی گردند. گمان میرود که مگسها نیز ممکن است با روح در حال خروج مردگان مرتبط بوده باشند، زیرا اغلب در نزدیکی اجساد یافت میشوند. در مصر مدرن، اعتقاد مشابهی در برخی مناطق وجود دارد که به مگس های سبز براق ضربه نزنید، زیرا ممکن است حامل روح یک فرد اخیراً فوت شده باشند. [94]
در یک افسانه یونانی که کمتر شناخته شده است ، یک دوشیزه بسیار پرحرف و پرحرف به نام میا (به معنی پرواز) الهه ماه سلن را با تلاش برای اغوای معشوقش، اندیمیون خفته ، خشمگین کرد و در نتیجه توسط الهه خشمگین به مگس تبدیل شد. که حالا به یاد زندگی گذشته اش همیشه خواب را از مردم سلب می کند. [95] [96] در Prometheus Bound که به نمایشنامهنویس تراژیک آتنی Aeschylus نسبت داده میشود ، مگس گاد که توسط هرا همسر زئوس فرستاده شده، معشوقهاش Io را که تبدیل به گاو شده است تعقیب و عذاب میدهد . چشمان گلهدار آرگوس : [97] [98] "ایو: آه! هه! دوباره خار، نیش مگس گاو! ای زمین، زمین، پنهان، شکل توخالی - آرگوس - آن چیز بد - صد- چشم." [98] ویلیام شکسپیر ، با الهام از آیسخولوس، تام اوبدلام را در شاه لیر دارد ، "که شیطان پلید او را از میان آتش و از میان شعله، از میان تندرو و گرداب، باتلاق و باتلاق هدایت کرده است." تعقیب [98] در آنتونی و کلئوپاترا ، شکسپیر به طور مشابه خروج عجولانه کلئوپاترا از میدان نبرد Actium را به گاوی تشبیه می کند که توسط یک مگس گاد تعقیب می شود. [99] اخیراً، در سال 1962، وینسنت دتیر، زیستشناس، مگس را بشناسید ، خوانندگان عمومی را با رفتار و فیزیولوژی مگس آشنا کرد. [100]
Musca depicta ("مگس نقاشی شده" در لاتین) به تصویر کشیدن یک مگس به عنوان عنصری نامحسوس از نقاشی های مختلف است. این ویژگی در نقاشی های قرن 15 و 16 بسیار رایج بود و وجود آن را می توان به دلایل مختلفی توضیح داد. [101]
مگس ها در فرهنگ عامه در مفاهیمی مانند ساخت مستند پرواز بر روی دیوار در تولید فیلم و تلویزیون ظاهر می شوند . این نام استعاری نشان میدهد که رویدادها به طور صریح دیده میشوند ، همانطور که مگس ممکن است آنها را ببیند. [102] مگس ها از طراحی روبات های پرنده مینیاتوری الهام گرفته اند. [103] فیلم پارک ژوراسیک استیون اسپیلبرگ در سال 1993 بر این ایده تکیه داشت که DNA را می توان در محتویات معده مگس خونخوار فسیل شده در کهربا حفظ کرد ، اگرچه مکانیسم توسط دانشمندان رد شده است. [104]
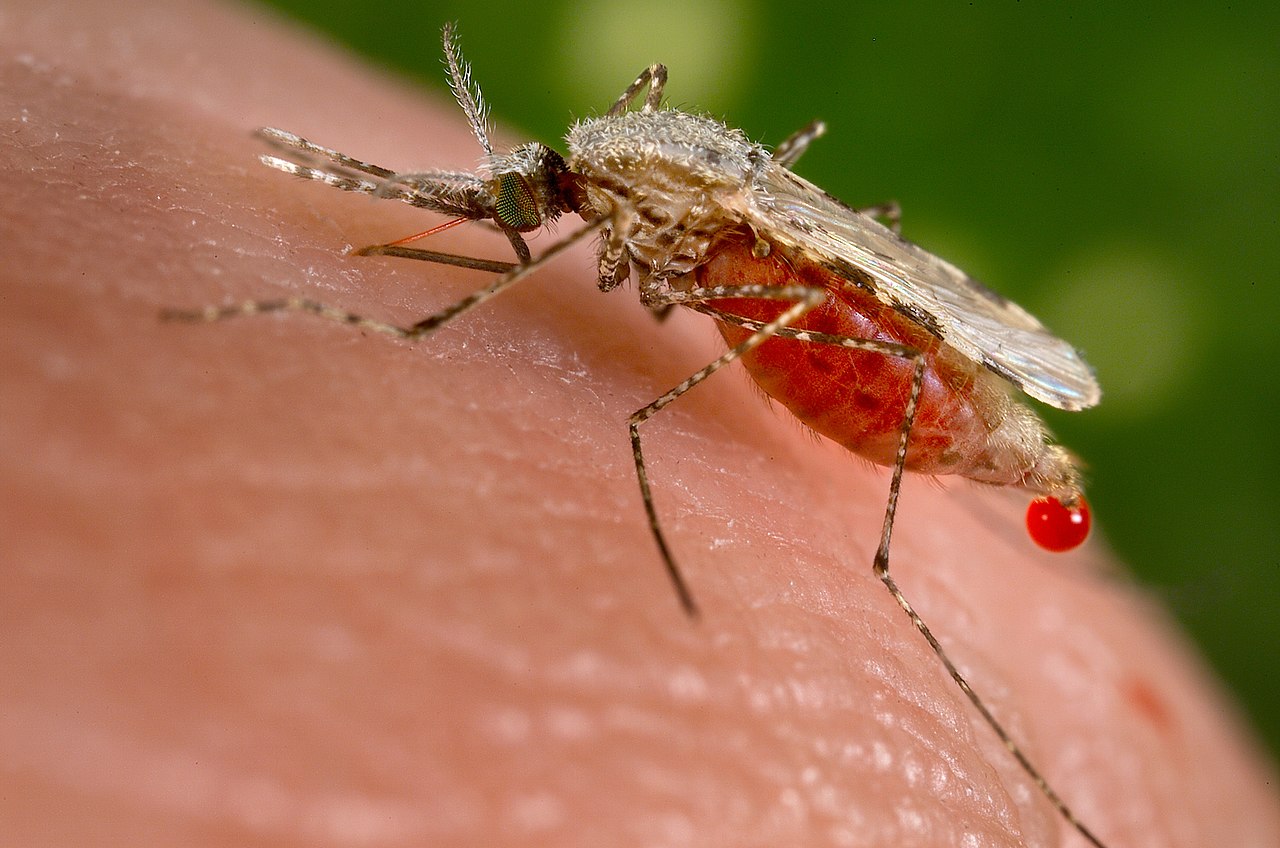
دیپتران ها گروه مهمی از حشرات هستند و تأثیر قابل توجهی بر محیط زیست دارند. برخی از مگس های معدنی برگ (Agromyzidae)، مگس میوه (Tephritidae و Drosophilidae) و میخ های صفراوی (Cecidomyiidae) آفات محصولات کشاورزی هستند. سایرین مانند مگس تسه ، کرم پیچ و مگس (Oestridae) به دام ها حمله می کنند، زخم ایجاد می کنند، بیماری را گسترش می دهند و آسیب اقتصادی قابل توجهی ایجاد می کنند. مقاله: مگس انگلی حیوانات اهلی را ببینید . تعداد کمی حتی می توانند باعث میاز در انسان شوند. برخی دیگر مانند پشهها (Culicidae)، مگسهای سیاه (Simuliidae) و مگسهای تخلیه (Psychodidae) بر سلامت انسان تأثیر میگذارند و به عنوان ناقل بیماریهای مهم گرمسیری عمل میکنند. در این میان پشه های آنوفل ناقل مالاریا ، فیلاریازیس و آربوویروس ها هستند . پشه های Aedes aegypti ناقل تب دنگی و ویروس زیکا هستند . مگس سیاه حامل کوری رودخانه ; پشه خاکی ناقل لیشمانیوز سایر دیپتران ها برای انسان آزاردهنده هستند، به خصوص زمانی که به تعداد زیاد وجود داشته باشند. اینها شامل مگس های خانگی است که غذا را آلوده می کند و بیماری های منتقله از طریق غذا را گسترش می دهد. پشه خاکی و پشه خاکی گزنده (Ceratopogonidae) و مگس خانگی و مگس پایدار (Muscidae). [34] در مناطق گرمسیری، مگسهای چشمی ( Chloropidae ) که در جستجوی مایعات به چشم میآیند، میتوانند در برخی فصول آزاردهنده باشند. [105]
بسیاری از دیپتران ها نقش هایی را ایفا می کنند که برای انسان مفید است. مگسهای خانگی، مگسهای بادکنکی و مگسهای قارچی (Mycetophilidae) لاشخور هستند و به تجزیه کمک میکنند. مگس های سارق (Asilidae)، تاچینیدها (Tachinidae) و مگس خنجر و مگس بادکنکی (Empididae) شکارچیان و انگل های دیگر حشرات هستند که به کنترل انواع آفات کمک می کنند. بسیاری از دیپتران ها مانند مگس زنبور عسل (Bombyliidae) و مگس شناور (Syrphidae) گرده افشان گیاهان زراعی هستند. [34]

مگس میوه مگس سرکهبه دلیل سهولت پرورش و پرورش آن در آزمایشگاه، ژنوم کوچک آن ، و این واقعیت که بسیاری از ژنهای آن در یوکاریوتهای بالاتر مشابه دارند، مدتهاست به عنوان یک ارگانیسم نمونه در تحقیقات مورد استفاده قرار میگیرد. . تعداد زیادی از مطالعات ژنتیکی بر اساس این گونه انجام شده است. اینها تأثیر عمیقی بر مطالعه بیان ژن ، مکانیسم های تنظیم کننده ژن و جهش داشته اند . مطالعات دیگر فیزیولوژی ، پاتوژنز میکروبی و توسعه را در میان سایر موضوعات تحقیقاتی بررسی کرده اند. [106] مطالعات روی روابط دوپترانه توسط ویلی هنیگ به توسعه cladistics کمک کرد ، تکنیکهایی که او برای شخصیتهای مورفولوژیکی به کار برد اما اکنون برای استفاده با توالیهای مولکولی در فیلوژنتیک سازگار شده است. [107]
حشره هایی که روی اجساد یافت می شوند برای حشره شناسان پزشکی قانونی مفید هستند . گونه های ماگوت را می توان با ویژگی های تشریحی و با تطبیق DNA آنها شناسایی کرد . حشرههای گونههای مختلف مگسها در زمانهای کاملاً مشخصی پس از مرگ قربانی از اجساد و لاشهها دیدن میکنند و شکارچیان آنها نیز مانند سوسکهای خانواده Histeridae . بنابراین، وجود یا عدم وجود گونه های خاص شواهدی را برای زمان پس از مرگ، و گاهی اوقات جزئیات دیگری مانند محل مرگ، زمانی که گونه ها در زیستگاه های خاصی مانند جنگل ها محدود می شوند، ارائه می دهد . [108]

برخی از گونههای حشرهها مانند لارو مگسهای بادکنک (آدمها) و لارو بطری آبی ( کاستر ) به صورت تجاری پرورش داده میشوند. آنها به عنوان طعمه در ماهیگیری و به عنوان غذا برای حیوانات گوشتخوار (که به عنوان حیوان خانگی، در باغ وحش ها یا برای تحقیق نگهداری می شوند) مانند برخی از پستانداران ، [109] ماهی ها ، خزندگان ، و پرندگان فروخته می شوند . پیشنهاد شده است که لارو مگس می تواند در مقیاس بزرگ به عنوان غذا برای مرغ های پرورشی، خوک ها و ماهی ها استفاده شود. با این حال، مصرف کنندگان با گنجاندن حشرات در غذای خود مخالف هستند و استفاده از حشرات در خوراک دام در مناطقی مانند اتحادیه اروپا غیرقانونی است . [110] [111]
از لارو مگس می توان به عنوان یک ابزار زیست پزشکی برای مراقبت و درمان زخم استفاده کرد. درمان دبریدمان حشره (MDT) استفاده از لارو مگس دمنده برای برداشتن بافت مرده از زخم است که معمولاً قطع عضو است. از لحاظ تاریخی، این برای قرنها، چه عمدی و چه غیرعمدی، در میدانهای جنگ و در بیمارستانهای اولیه مورد استفاده قرار گرفته است. [112] برداشتن بافت مرده باعث رشد سلولی و بهبود زخم سالم می شود. لاروها همچنین دارای خواص بیوشیمیایی مانند فعالیت ضد باکتریایی هستند که در ترشحات آنها هنگام تغذیه یافت می شود. [113] این ماگت های دارویی یک درمان ایمن و مؤثر برای زخم های مزمن هستند. [114]
پنیر ساردینیا casu marzu در معرض مگس هایی قرار می گیرد که به عنوان کاپیتان پنیر شناخته می شوند مانند Piophila casei ، اعضای خانواده Piophilidae . [115] فعالیت های گوارشی لارو مگس پنیر را نرم می کند و عطر را به عنوان بخشی از فرآیند بلوغ تغییر می دهد. زمانی مقامات اتحادیه اروپا فروش پنیر را ممنوع کردند و یافتن آن سخت می شد، [116] اما این ممنوعیت به این دلیل برداشته شد که پنیر یک محصول سنتی محلی است که با روش های سنتی تهیه می شود. [117]
مگس ها یک خطر برای سلامتی هستند و به دلیل بویی که دارند جذب توالت می شوند. مجله New Scientist تله ای برای این مگس ها پیشنهاد کرد. لوله ای که به عنوان دودکش عمل می کند به توالت تعبیه شده بود که مقداری نور را برای جذب این مگس ها تا انتهای این لوله می داد، جایی که یک پوشش گاز مانع از فرار به هوای بیرون می شد به طوری که آنها به دام افتادند و مردند. توالت ها به طور کلی داخل تاریک هستند، به خصوص اگر در بسته باشد.
{{cite book}}: CS1 maint: multiple names: authors list (link)تقریباً هر عنصر در جهان ممکن است شخصی باشد، و حتی کوچکترین آنها مانند سنجاب کوچک و آن حشرات کوچک یاوران و مربیان خدا و انسان در اسطوره ها، مگس بزرگ (
Dǫ'soh
) و Ripener (سوسک ذرت) ('
Anilt'ánii 'At'ééd
) (Wyman and Bailey 1964:29-30, 51, 137-144)، به اندازه خورشید بزرگ برای تعادل هماهنگ جهان ضروری هستند.
ژنرال
آناتومی
توصیف کنندگان