Ctenophora ( / t ə ˈ n ɒ f ər ə / tə- NOF -ər-ə ؛ sg . : ctenophore / ˈ t ɛ n ə f ɔːr , ˈ t iː n ə -/ TEN -ə- برای ، ; آنها به دلیل گروه های مژک که برای شنا استفاده می کنند (که معمولاً به آنها "شانه" گفته می شود) قابل توجه هستند و آنها بزرگترین حیواناتی هستند که با کمک مژه ها شنا می کنند.
بسته به گونه، اندازه ctenophoreهای بالغ از چند میلی متر تا 1.5 متر (5 فوت) متغیر است. در حال حاضر تنها 186 گونه زنده شناخته شده است. [7]
بدن آنها از توده ای از ژله تشکیل شده است که لایه ای به ضخامت دو سلول در خارج و لایه ای دیگر حفره داخلی را پوشانده است. این شاخه دارای طیف وسیعی از فرمهای بدنی است، از جمله سایدیپیدهای تخممرغی با یک جفت شاخک جمع شونده که طعمه را میگیرند، پلاتیکتنیدهای صاف بهطور کلی بدون شانه، و بروئیدهای دهان درشت ، که طعمههای تنوفورهای دیگر میشوند.
تقریباً همه ctenophore ها به عنوان شکارچی عمل می کنند و طعمه را از لاروهای میکروسکوپی و روتیفرها گرفته تا بالغ سخت پوستان کوچک می گیرند . استثناء بچه های دو گونه است که به عنوان انگل روی نمک هایی که بزرگسالان گونه خود از آنها تغذیه می کنند زندگی می کنند.
با وجود بدن نرم و ژلاتینیشان، فسیلهایی که تصور میشود نمایانگر کتنوفورها هستند، در لاگرستاتن ظاهر میشوند که قدمت آن به اوایل کامبرین ، حدود 525 میلیون سال پیش بازمیگردد. موقعیت ctenophores در "درخت زندگی" مدتهاست در مطالعات فیلوژنتیک مولکولی مورد بحث بوده است. زیست شناسان پیشنهاد کردند که کتنوفورها دومین اصل و نسب حیوانات منشعب اولیه را تشکیل می دهند و اسفنج ها گروه خواهر همه حیوانات چند سلولی دیگر هستند ( فرضیه خواهر پوریفرا ). [8] سایر زیست شناسان ادعا می کنند که ctenophore ها زودتر از اسفنج ها پدید آمدند ( فرضیه خواهر Ctenophora )، که خود قبل از انشعاب بین cnidarians و دولاتری ها ظاهر شدند . [9] [10] پیسانی و همکاران . دادهها را دوباره تجزیه و تحلیل کرد و پیشنهاد کرد که الگوریتمهای کامپیوتری مورد استفاده برای تجزیه و تحلیل با وجود ژنهای ctenophore خاص که به طور قابلتوجهی با ژنهای دیگر گونهها متفاوت بودند، گمراه شدند. [11] [12] [ صفحه مورد نیاز ] تجزیه و تحلیل پیگیری توسط Whelan و همکاران. (2017) [13] از فرضیه 'خواهر Ctenophora' پشتیبانی بیشتری کرد. این موضوع همچنان یک موضوع اختلاف طبقه بندی است. [14] [15] شولتز و همکاران. (2023) تغییرات غیرقابل برگشتی را در synteny در خواهر Ctenophora، Myriazoa ، متشکل از بقیه حیوانات یافت. [16]

در میان فیلاهای جانوری، Ctenophores پیچیدهتر از اسفنجها ، تقریباً به پیچیدگی cnidarians ( ژلهماهی ، شقایقهای دریایی و غیره) و پیچیدهتر از دولایهها (که تقریباً همه حیوانات دیگر را شامل میشود) هستند. بر خلاف اسفنج ها، هم ctenophores و هم cnidarians دارای موارد زیر هستند:
Ctenophore ها با داشتن کلوبلاست هایی که چسبنده هستند و به طعمه می چسبند از همه حیوانات دیگر متمایز می شوند ، اگرچه تعداد کمی از گونه های ctenophore فاقد آنها هستند. [18] [19]
مانند cnidarians، ctenophore ها دارای دو لایه اصلی سلول هستند که یک لایه میانی از مواد ژله مانند را ساندویچ می کنند، که در cnidarians و ctenophores مزوگلیا نامیده می شود. حیوانات پیچیده تر دارای سه لایه سلولی اصلی و بدون لایه ژله مانند هستند. از این رو ctenophores و cnidarians به طور سنتی برچسب diploblastic نامیده می شوند . [18] [20] هر دو ctenophores و cnidarians یک نوع ماهیچه دارند که در حیوانات پیچیدهتر، از لایه سلول میانی ناشی میشود ، [21] و در نتیجه برخی از کتابهای درسی اخیر ctenophores را به عنوان تریپلوبلاستیک طبقهبندی میکنند ، [22] در حالی که دیگران هنوز آنها را به عنوان دیپلوپلاستیک در نظر می گیرند. [۱۸] ژلههای شانهای بیش از ۸۰ نوع سلول مختلف دارند ، که از تعداد گروههای دیگر مانند پلاکوزوئنها، اسفنجها، cnidarians و برخی دوجانبههای عمیقشاخه بیشتر است. [23]
اندازه آنها از حدود 1 میلی متر (0.04 اینچ) تا 1.5 متر (5 فوت) است، [22] [24] ctenophore ها بزرگترین حیوانات غیر استعماری هستند که از مژگان ("مو") به عنوان روش اصلی حرکت خود استفاده می کنند. [22] بیشتر گونهها دارای هشت نوار به نام ردیفهای شانهای هستند که طول بدنشان را میکشد و نوارهایی از مژکهای شانهمانند به نام «ctenes» دارند که در امتداد ردیفهای شانه روی هم قرار میگیرند به طوری که وقتی مژکها میکوبند، مژکهای هر شانه روی هم قرار میگیرند. شانه زیر را لمس کنید. [22] نام "ctenophora" به معنای "شانه دار"، از یونانی κτείς (شکل ساقه κτεν- ) به معنای "شانه" و پسوند یونانی -φορος به معنای "حمل" است. [25]
برای یک شاخه با گونه های نسبتاً کمی، ctenophore ها دارای طیف گسترده ای از طرح های بدن هستند. [22] گونه های ساحلی باید به اندازه کافی سخت باشند تا بتوانند امواج و ذرات رسوبی چرخان را تحمل کنند، در حالی که برخی از گونه های اقیانوسی آنقدر شکننده هستند که گرفتن دست نخورده آنها برای مطالعه بسیار دشوار است. [19] علاوه بر این، گونههای اقیانوسی به خوبی حفظ نمیشوند، [19] و عمدتاً از عکسها و یادداشتهای ناظران شناخته میشوند. [30] از این رو بیشترین توجه تا همین اواخر بر روی سه جنس ساحلی - Pleurobrachia ، Beroe و Mnemiopsis متمرکز بود . [19] [31] حداقل دو کتاب درسی توصیفات خود را از ctenophores بر اساس سیدیپیپ Pleurobrachia قرار داده اند . [18] [22]
از آنجایی که بدن بسیاری از گونه ها تقریباً به صورت شعاعی متقارن است ، محور اصلی دهان به آبورال (از دهان تا انتهای مخالف) است. با این حال، از آنجایی که تنها دو کانال در نزدیکی استاتوسیست به منافذ مقعد ختم میشوند ، کتنوفورها هیچ تقارن آینهای ندارند، اگرچه بسیاری از آنها دارای تقارن چرخشی هستند. به عبارت دیگر، اگر حیوان در یک نیم دایره بچرخد، مانند زمانی است که شروع کرده است. [32]
شاخه Ctenophore دارای طیف گسترده ای از اشکال بدن است، از جمله پلاتیکتنیدهای پهن شده و در اعماق دریا ، که در آن حشرات بالغ اکثر گونه ها فاقد شانه هستند، و beroids ساحلی ، که فاقد شاخک هستند و با استفاده از دهان های بزرگ مسلح به گروه ها، سایر ctenophore ها را شکار می کنند. مژک های بزرگ و سفت شده که به عنوان دندان عمل می کنند.

بدن کتنوفورها مانند چتر دریایی ، شقایقهای دریایی و غیره، از یک مزوگلیای نسبتاً ضخیم و ژلهمانند تشکیل شده است که بین دو اپیتلیوم ، لایههایی از سلولها که توسط اتصالات بین سلولی و توسط یک غشای پایه فیبری محدود شدهاند، قرار گرفته است . ترشح کردن . [18] [22] اپیتلیوم ctenophores دارای دو لایه سلولی به جای یک لایه است و برخی از سلول های لایه بالایی دارای چندین مژک در هر سلول هستند. [22]
لایه بیرونی اپیدرم ( پوست خارجی) از: سلول های حسی تشکیل شده است. سلول هایی که مخاط ترشح می کنند که از بدن محافظت می کند. و سلول های بینابینی که می توانند به انواع دیگر سلول تبدیل شوند. در قسمتهای تخصصی بدن، لایه بیرونی همچنین حاوی کلوبلاستهایی است که در امتداد سطح شاخکها یافت میشوند و برای گرفتن طعمه یا سلولهایی که چندین مژک بزرگ دارند برای حرکت استفاده میشوند. لایه داخلی اپیدرم شامل یک شبکه عصبی و سلول های میواپیتلیال است که به عنوان ماهیچه عمل می کنند . [22]
حفره داخلی شکل می گیرد: دهانی که معمولاً می تواند توسط ماهیچه ها بسته شود. حلق ("گلو" ) ؛ ناحیه وسیع تری در مرکز که به عنوان معده عمل می کند . و سیستم کانال های داخلی. این ها از طریق مزوگلیا به فعال ترین قسمت های حیوان منشعب می شوند: دهان و حلق. ریشه شاخک ها، در صورت وجود؛ تمام قسمت زیرین هر ردیف شانه؛ و چهار شاخه در اطراف مجتمع حسی در انتهای دور دهان - دو شاخه از این چهار شاخه به منافذ مقعد ختم می شوند. سطح داخلی حفره با یک اپیتلیوم به نام گاسترودرم پوشیده شده است . دهان و حلق هر دو مژک و ماهیچه های توسعه یافته دارند. در سایر قسمتهای سیستم کانال، گاسترودرم در نزدیکترین و دورترین طرفها از اندامی که تامین میکند متفاوت است. سمت نزدیکتر از سلولهای مغذی بلندی تشکیل شده است که مواد مغذی را در واکوئلها (بخشهای داخلی) ذخیره میکنند، سلولهای زایا که تخمک یا اسپرم تولید میکنند و فوتوسیتهایی که بیولومینسانس تولید میکنند . دورترین سمت از اندام با سلول های مژک دار پوشیده شده است که آب را از طریق کانال ها به گردش در می آورند، منافذی که توسط حلقه های مژک دوتایی احاطه شده اند و به مزوگلیا متصل می شوند. [22]
هنگامی که طعمه بلعیده می شود، توسط آنزیم ها و انقباضات عضلانی حلق در حلق مایع می شود . دوغاب حاصل با ضربه مژه ها از طریق سیستم کانال پخش می شود و توسط سلول های تغذیه کننده هضم می شود. گل های مژگانی در کانال ها ممکن است به انتقال مواد مغذی به ماهیچه های مزوگلیا کمک کنند. منافذ مقعد ممکن است ذرات کوچک ناخواسته را خارج کنند، اما بیشتر مواد ناخواسته از طریق دهان بازگردانده می شوند. [22]
اطلاعات کمی در مورد چگونگی خلاص شدن کتنوفورها از مواد زائد تولید شده توسط سلول ها وجود دارد. روزت های مژگانی موجود در گاسترودرم ممکن است به حذف مواد زائد از مزوگلیا کمک کند و همچنین ممکن است با پمپاژ آب به داخل یا خارج از مزوگلیا به تنظیم شناوری حیوان کمک کند . [22]
سطح بیرونی معمولاً دارای هشت ردیف شانه به نام صفحات شنا است که برای شنا استفاده می شود. ردیفها به گونهای هستند که از نزدیک دهان ("قطب دهان") به سمت مخالف ("قطب آبورال") کشیده شدهاند، و کم و بیش به طور مساوی در اطراف بدن قرار گرفتهاند، [18] اگرچه الگوهای فاصله براساس گونهها و گونهها متفاوت است. در بیشتر گونهها، ردیفهای شانهها تنها بخشی از فاصله قطب آبورال را به سمت دهان گسترش میدهند. «شانهها» (همچنین «ctenes» یا «صفحات شانهای» نامیده میشوند) در هر ردیف قرار دارند و هر کدام از هزاران مژک بلند غیرعادی تا 2 میلیمتر (0.08 اینچ) تشکیل شده است. بر خلاف مژکها و تاژکهای معمولی که دارای ساختار رشتهای هستند که در الگوی 9 + 2 مرتب شدهاند، این مژکها در یک الگوی 9 + 3 قرار گرفتهاند، جایی که مشکوک است که رشته فشرده اضافی عملکرد حمایتی داشته باشد. [33] اینها معمولاً طوری می زنند که حرکت پیشرانه از دهان دور می شود، اگرچه می توانند جهت را نیز معکوس کنند. از این رو کتنوفورها بر خلاف چتر دریایی معمولاً در جهتی شنا می کنند که دهان در حال غذا خوردن است . [22] هنگام تلاش برای فرار از شکارچیان، یک گونه می تواند تا شش برابر سرعت عادی خود شتاب بگیرد. [34] برخی از گونههای دیگر بهعنوان بخشی از رفتار گریز خود، با معکوس کردن حرکت قدرتی مژکهای صفحه شانه، جهت معکوس میکنند.
مشخص نیست که چگونه ctenophore ها شناوری خود را کنترل می کنند، اما آزمایشات نشان داده است که برخی از گونه ها برای انطباق با آب با چگالی های مختلف به فشار اسمزی متکی هستند. [35] مایعات بدن آنها معمولاً به اندازه آب دریا غلیظ است . اگر آنها وارد آب شور تراکم کمتری شوند، روزت های مژگانی در حفره بدن ممکن است این را به داخل مزوگلیا پمپ کنند تا حجم آن را افزایش دهند و تراکم آن را کاهش دهند تا از غرق شدن جلوگیری کنند. برعکس، اگر آنها از آب شور به آب دریا با قدرت کامل حرکت کنند، ممکن است روزت ها آب را از مزوگلیا برای کاهش حجم و افزایش چگالی آن به بیرون پمپاژ کنند. [22]
Ctenophore ها مغز یا سیستم عصبی مرکزی ندارند ، اما در عوض دارای یک شبکه عصبی زیر اپیدرمی هستند (به جای تار عنکبوت) که حلقه ای دور دهان تشکیل می دهد و متراکم ترین نزدیک به ساختارهایی مانند ردیف شانه، حلق، شاخک ها (در صورت وجود) و حسی است. پیچیده ترین دورتر از دهان [22] ارتباط بین سلول های عصبی از دو روش مختلف استفاده می کند. مشخص شده است که برخی از نورونها دارای اتصالات سیناپسی هستند ، اما نورونهای موجود در شبکه عصبی بهجای اتصال توسط سیناپسها، به دلیل ادغام در یک سینسیتیوم بسیار متمایز هستند . برخی از جانوران خارج از ctenophore ها نیز دارای سلول های عصبی ذوب شده هستند، اما هرگز به اندازه ای که یک شبکه عصبی کامل را تشکیل دهند. [36] [37] [38] فسیلها نشان میدهند که گونههای کامبرین سیستم عصبی پیچیدهتری داشتند، با اعصاب بلند که با حلقهای در اطراف دهان به هم متصل میشدند. تنها کتنوفور شناخته شده با اعصاب بلند امروزه Euplokamis از راسته Cydippida است. [39] سلولهای عصبی آنها از همان سلولهای پیشساز ایجاد میشوند که کلوبلاستها. [40]
علاوه بر این، یک شبکه عصبی مزوگلیال کمتر سازمان یافته متشکل از نوریت های منفرد وجود دارد. بزرگترین ویژگی حسی منفرد، اندام آبورال (در انتهای مخالف دهان) است که با شبکه عصبی خودش زیر آن خط کشیده شده است. [41] جزء اصلی این اندام یک استاتوسیست است ، یک حسگر تعادل متشکل از یک استاتولیت، یک دانه ریز کربنات کلسیم، که بر روی چهار دسته مژک ، به نام "تعادل کننده"، که جهت آن را حس می کند، حمایت می شود. استاتوسیست توسط یک گنبد شفاف ساخته شده از مژک های بلند و بی حرکت محافظت می شود. یک ctenophore به طور خودکار سعی نمی کند استاتولیت را به طور یکسان روی همه متعادل کننده ها قرار دهد. در عوض، پاسخ آن توسط "خلق" حیوان، به عبارت دیگر، وضعیت کلی سیستم عصبی تعیین می شود. به عنوان مثال، اگر یک تنوفور با شاخک های دنباله دار شکار را بگیرد، اغلب چند ردیف شانه را به سمت عقب قرار می دهد و دهان را به سمت طعمه می چرخاند. [22]
تحقیقات از این فرضیه پشتیبانی می کند که لاروهای مژک دار در cnidarians و biterians منشأ باستانی و مشترک دارند. [42] اندام آپیکال لارو در تشکیل سیستم عصبی نقش دارد. [43] اندام ابورال ژله های شانه ای با اندام آپیکال در حیوانات دیگر همولوگ نیست و بنابراین تشکیل سیستم عصبی آنها منشا جنینی متفاوتی دارد. [44]
سلول های عصبی Ctenophore و سیستم عصبی در مقایسه با سایر حیوانات، بیوشیمی متفاوتی دارند. برای مثال، آنها فاقد ژنها و آنزیمهای مورد نیاز برای ساخت انتقالدهندههای عصبی مانند سروتونین ، دوپامین ، اکسید نیتریک ، اکتوپامین ، نورآدرنالین و غیره هستند که در غیر این صورت در همه حیوانات دیگر با سیستم عصبی دیده میشود، با ژنهایی که برای گیرندههای هر یک از آنها کد میکنند. انتقال دهنده های عصبی از دست رفته است [45] کاتالاز تک عملکردی (CAT)، یکی از سه خانواده اصلی آنزیم های آنتی اکسیدانی که پراکسید هیدروژن را هدف قرار می دهد ، یک مولکول سیگنال دهنده مهم برای فعالیت سیناپسی و عصبی، نیز به احتمال زیاد به دلیل از دست دادن ژن وجود ندارد. [46] مشخص شده است که آنها از ال-گلوتامات به عنوان یک انتقال دهنده عصبی استفاده می کنند و دارای تنوع غیرعادی بالایی از گیرنده های گلوتامات یونوتروپیک و ژن برای سنتز و انتقال گلوتامات در مقایسه با سایر متازوئن ها هستند. [47] محتوای ژنومی ژن های سیستم عصبی کوچکترین موجود در بین حیوانات است و می تواند حداقل نیازهای ژنتیکی برای یک سیستم عصبی عملکردی را نشان دهد. [48] این واقعیت که بخشهایی از سیستم عصبی دارای نورونهای ذوب شده مستقیم، بدون سیناپس هستند، نشان میدهد که کتنوفورها ممکن است یک گروه خواهر با متازوئنهای دیگر تشکیل دهند، که به طور مستقل یک سیستم عصبی را توسعه دادهاند. [38] اگر کتنوفورها گروه خواهر همه متازوئنهای دیگر باشند، سیستم عصبی ممکن است در اسفنجها و پلاکوزوئنها از بین رفته باشد یا بیش از یک بار در میان متازوئنها بوجود آمده باشد. [49]

تنوفورهای سیدیپیدی دارای بدنه هایی هستند که کم و بیش گرد، گاهی تقریباً کروی و گاهی اوقات بیشتر استوانه ای یا تخم مرغی شکل هستند. انگور فرنگی معمولی ساحلی، Pleurobrachia ، گاهی اوقات دارای بدنی تخم مرغی شکل با دهان در انتهای باریک است، [22] اگرچه برخی از افراد به طور یکنواخت گرد هستند. از طرف مقابل بدن یک جفت شاخک بلند و باریک بیرون میآید که هر کدام در غلافی قرار دارند که میتوان آن را داخل آن کشید. [18] برخی از گونههای سیدیپیدها دارای بدنهای هستند که به میزانهای مختلف صاف شدهاند به طوری که در صفحه شاخکها پهنتر هستند. [22]
شاخکهای ctenophores cydippid معمولاً با شاخکهای کوچک ("شاخکهای کوچک") حاشیهدار هستند، اگرچه تعدادی از جنسها شاخکهای ساده بدون این شاخههای جانبی دارند. شاخک ها و شاخک ها به طور متراکمی با کلوبلاست های میکروسکوپی پوشیده شده اند که طعمه را با چسبیدن به آن می گیرند. کلوبلاست ها سلول های قارچی شکل تخصصی در لایه بیرونی اپیدرم هستند و دارای سه جزء اصلی هستند: یک سر گنبدی با وزیکول ها (حفره هایی) که حاوی چسب هستند. ساقه ای که سلول را در لایه زیرین اپیدرم یا در مزوگلیا لنگر می اندازد. و یک نخ مارپیچ که دور ساقه می پیچد و به سر و به ریشه ساقه می چسبد. عملکرد نخ مارپیچ نامشخص است، اما ممکن است زمانی که طعمه سعی در فرار دارد، استرس را جذب کند و در نتیجه از پاره شدن کلوبلاست جلوگیری کند. [22] یک گونه، Minictena luteola، که فقط 1.5 میلی متر قطر دارد، دارای پنج نوع مختلف سلول کلوبلاست است. [50] [51]
علاوه بر کلوبلاستها، اعضای جنس Haeckelia که عمدتاً از چتر دریایی تغذیه میکنند، نماتوسیتهای نیشزن قربانیان خود را در شاخکهای خود قرار میدهند - برخی از شاخههای برهنهخوار cnidaria به طور مشابه نماتوسیتها را برای دفاع در بدن خود وارد میکنند. [52] تنتیل Euplokamis به طور قابل توجهی با سایر cydippids متفاوت است: آنها حاوی ماهیچه مخطط هستند ، یک نوع سلول که در غیر این صورت در شاخه Ctenophora ناشناخته است. و زمانی که شل میشوند پیچ میخورند، در حالی که تنتیلای همه کتنوفورهای شناختهشده دیگر وقتی شل میشوند کشیده میشوند. شاخک Euplokamis دارای سه نوع حرکت است که برای گرفتن طعمه استفاده می شود: آنها ممکن است خیلی سریع (در 40 تا 60 میلی ثانیه ) خارج شوند. آنها می توانند چروکیده شوند، که ممکن است با رفتاری شبیه کرم های پلانکتونیک طعمه را فریب دهند. و شکار گرد را می پیچند. تلنگر منحصر به فرد یک حرکت باز شدن پیچ است که توسط انقباض عضله مخطط انجام می شود . حرکت چرخشی توسط عضلات صاف ایجاد می شود ، اما از نوع بسیار تخصصی است. حلقه زدن به دور طعمه عمدتاً با بازگشت تنتیل به حالت غیرفعال خود انجام می شود، اما ممکن است سیم پیچ ها توسط ماهیچه صاف سفت شوند. [53]
هشت ردیف شانه وجود دارد که از نزدیک دهان تا انتهای مخالف کشیده شده اند و به طور مساوی دور بدن قرار گرفته اند. [18] "شانه ها" در ریتم متاکرونال و نه مانند یک موج مکزیکی می زند . [54] از هر متعادل کننده در استاتوسیست یک شیار مژگانی از زیر گنبد خارج می شود و سپس برای اتصال به دو ردیف شانه مجاور جدا می شود و در برخی گونه ها در امتداد ردیف های شانه کشیده می شود. این یک سیستم مکانیکی برای انتقال ریتم ضربان از شانه ها به متعادل کننده ها از طریق اختلالات آب ایجاد شده توسط مژک ها تشکیل می دهد. [55]
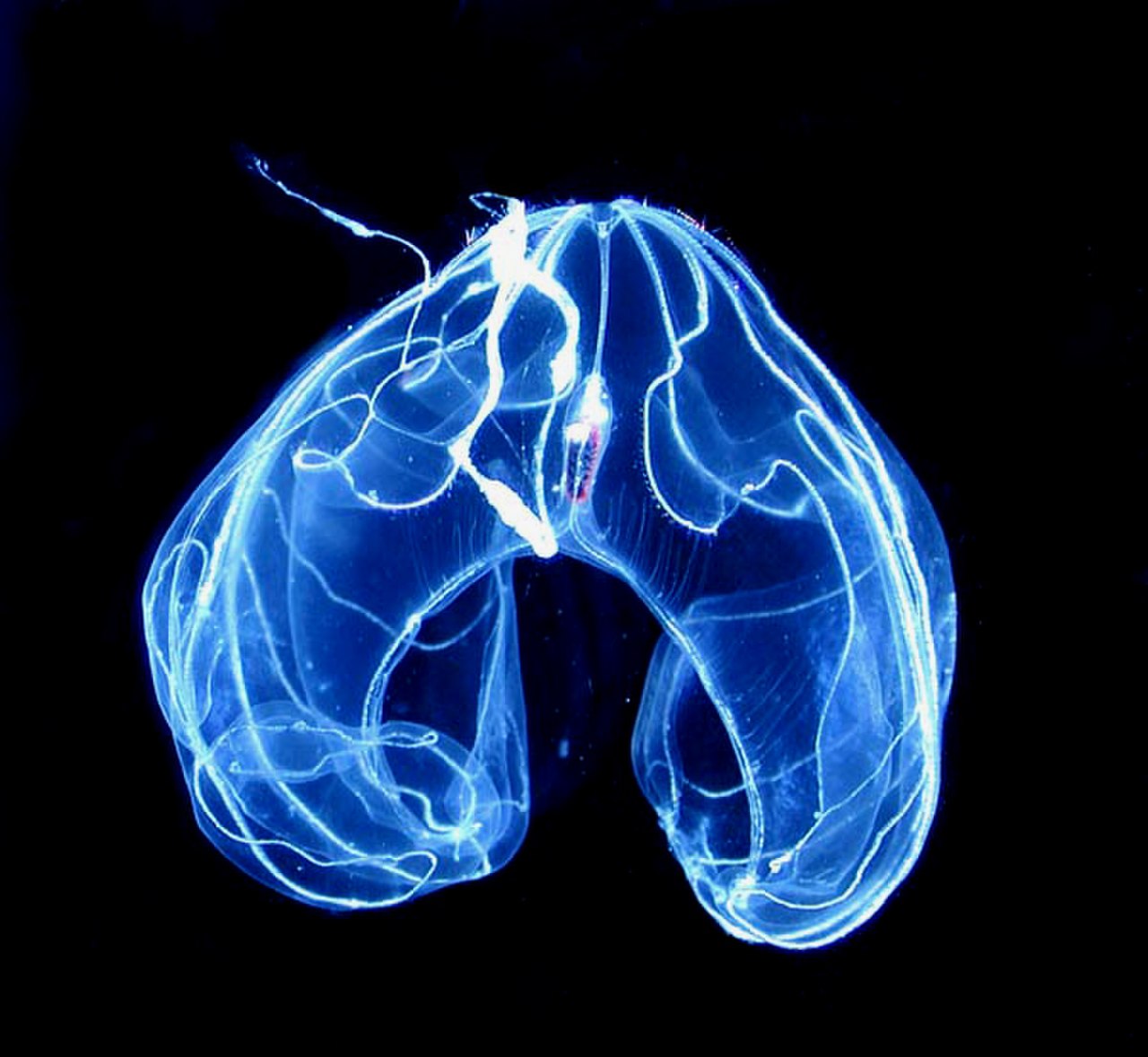
لوباتا دارای یک جفت لوب است که امتدادهای عضلانی و فنجانی بدن هستند که فراتر از دهان بیرون می آیند . شاخکهای نامحسوس آنها از گوشههای دهان سرچشمه میگیرند، در شیارهای پیچخورده میروند و روی سطح داخلی لوبها پخش میشوند (بهجای اینکه خیلی عقبتر مانند سیدیپیدا باشند). بین لوبهای دو طرف دهان، بسیاری از گونههای لوبات دارای چهار گوش، برآمدگیهای ژلاتینی با لبههای مژک هستند که جریانهای آبی تولید میکنند که به هدایت طعمه میکروسکوپی به سمت دهان کمک میکند. این ترکیب از ساختارها، لوبات ها را قادر می سازد تا به طور مداوم از طعمه های پلانکتون معلق تغذیه کنند . [22]
لوبات ها دارای هشت ردیف شانه هستند که از قطب آبورال منشا می گیرند و معمولاً از بدن تا لوب ها امتداد نمی یابند. در گونههای دارای (چهار) گوش، مژکهایی که لبههای گوشها را میگیرند، امتداد مژکها در چهار ردیف از ردیفهای شانهای هستند. اکثر لوبات ها هنگام حرکت در آب کاملاً منفعل هستند و از مژک های روی ردیف شانه های خود برای پیشرانه استفاده می کنند، [22] اگرچه لوکوته گوش های بلند و فعالی دارد که حرکات آنها نیز به پیشرانش کمک می کند. اعضای جنس لوبات Bathocyroe و Ocyropsis می توانند با دست زدن به لوب خود از خطر فرار کنند، به طوری که جت آب بیرون رانده شده آنها را خیلی سریع به عقب می راند. [56] برخلاف سیدیپیدها، حرکات شانههای لوباتها توسط اعصاب هماهنگ میشوند تا با اختلالات آب که توسط مژکها ایجاد میشوند، اما شانههای روی همان ردیف به همان سبک موج مکزیکی مانند ردیفهای شانهای که از نظر مکانیکی هماهنگ شدهاند سیدیپیدها و بیرویدها میکوبند. [55] این ممکن است باعث شود که لوبات ها بزرگتر از سیدیپیدها رشد کنند و شکل های کمتری شبیه تخم داشته باشند. [54]
گونه غیرمعمولی که برای اولین بار در سال 2000 توصیف شد، Lobatolampea tetragona ، به عنوان یک لوب طبقه بندی شده است، اگرچه لوب ها "اولیه" هستند و بدن در حالت شناور مانند مدوسا و هنگام استراحت در بستر دریا مانند دیسک است. [30]

بروئیدا که به نام نودا نیز شناخته می شود ، هیچ زائده تغذیه ای ندارد، اما حلق بزرگ آنها ، درست در داخل دهان بزرگ و پر کردن قسمت اعظم بدن کیسه مانند، دارای "ماکروسیلیا" در انتهای دهان است. این دستههای چند هزار مژک بزرگ میتوانند تکههای طعمهای را که برای بلع کامل آنقدر بزرگ هستند - تقریباً همیشه سایر کتنوفورها - «نیش بزنند». [57] در جلوی مزرعه ماکروسیلیا، روی دهان "لب" در برخی از گونههای Beroe ، یک جفت نوار باریک از سلولهای اپیتلیال چسبنده روی دیواره معده وجود دارد که دهان را هنگامی که حیوان تغذیه نمیکند، "زیپ" میبندد. ، با ایجاد اتصالات بین سلولی با نوار چسب مخالف. این بسته محکم جلوی حیوان را هنگامی که در حال تعقیب طعمه است ساده می کند . [58]
گانشیدا دارای یک جفت لوب کوچک دهان و یک جفت شاخک است . سطح مقطع بدن به جای بیضی شکل است و حلق بر روی سطوح داخلی لوب ها گسترش می یابد. [22]
Thalassocalycida ، تنها در سال 1978 کشف شد و تنها از یک گونه شناخته شد، [ 59] شبیه مدوزا هستند، با بدنهایی که در جهت دهانی-ابورال کوتاه شدهاند، و ردیفهای شانهای کوتاه در دورترین سطح از دهان، که از نزدیک منشا میگیرند. قطب آبورال آنها طعمه را با حرکات زنگ و احتمالاً با استفاده از دو شاخک کوتاه می گیرند. [22]
Cestida ("حیوانات کمربند") حیوانات پلانکتون نواری شکل هستند که دهان و اندام ابورال در وسط لبه های مخالف نوار قرار دارند . یک جفت ردیف شانه در امتداد هر لبه آبورال وجود دارد و شاخک از یک شیار در امتداد لبه دهانی بیرون میآید که در بیشتر سطح بال مانند بدن به عقب برمیگردد. سیستیدها می توانند با موج دار کردن بدن خود و همچنین با زدن ردیف شانه هایشان شنا کنند. دو گونه شناخته شده وجود دارد که در سرتاسر جهان در آب های گرم و معتدل پراکنده شده اند: Cestum veneris (" کمربند ناهید ") یکی از بزرگترین تنوفورها است - تا 1.5 متر (4.9 فوت) طول دارد و می تواند به آرامی یا بسیار سریع موج بزند. . موازی Velamen ، که معمولا کمتر از 20 سانتی متر (0.66 فوت) طول دارد، می تواند در آنچه به عنوان "حرکت دارتینگ" توصیف شده است، بسیار سریعتر حرکت کند. [22] [60]
اکثر Platyctenida دارای بدن های بیضی شکل هستند که در جهت دهانی- ابورال مسطح هستند، با یک جفت شاخک حامل شاخک در سطح ابورال. آنها با برگرداندن حلق و استفاده از آن به عنوان یک "پا" عضلانی به سطوح میچسبند و میخزند. همه به جز یکی از گونه های پلاتیکتنید شناخته شده فاقد ردیف شانه هستند. [22] پلاتیکتنیدها معمولاً رنگهای رمزآلود دارند، روی صخرهها، جلبکها یا سطوح بدن سایر بیمهرگان زندگی میکنند، و اغلب توسط شاخکهای بلندشان با شاخههای جانبی زیادی آشکار میشوند که از پشت کتنوفور به جریان میآیند.

بزرگسالان بیشتر گونهها میتوانند بافتهایی را که آسیب دیده یا برداشته شدهاند بازسازی کنند، [61] اگرچه فقط پلاتیکتنیدها با شبیهسازی تکثیر میشوند و از لبههای بدن صاف خود تکههایی میشکنند که به افراد جدید تبدیل میشوند. [22] تحقیقات آزمایشگاهی روی Mnemiopsis leidyi همچنین نشان میدهد که وقتی دو فرد قسمتهایی از بدنشان را جدا میکنند، میتوانند با هم ترکیب شوند، از جمله سیستم عصبی و گوارشیشان، حتی زمانی که این دو فرد از نظر ژنتیکی متفاوت هستند. پدیده ای که تاکنون فقط در ژله های شانه ای دیده شده است. [62]
آخرین جد مشترک (LCA) ctenophores هرمافرودیت بود . [63] برخی هرمافرودیتهای همزمان هستند که میتوانند همزمان تخمک و اسپرم تولید کنند، در حالی که برخی دیگر هرمافرودیتهای متوالی هستند که در آنها تخمک و اسپرم در زمانهای مختلف بالغ میشوند. دگردیسی وجود ندارد . [64] حداقل سه گونه شناخته شده است که جنس های جداگانه تکامل یافته اند ( دیواسی ). Ocyropsis crystallina و Ocyropsis maculata در جنس Ocyropsis و Bathocyroe fosteri از جنس Bathocyroe . [65] غدد جنسی در قسمتهایی از شبکه کانال داخلی زیر ردیفهای شانه قرار دارند و تخمکها و اسپرمها از طریق منافذ در اپیدرم آزاد میشوند. لقاح عموماً خارجی است ، اما پلاتیکتنیدها از لقاح داخلی استفاده میکنند و تخمها را تا زمانی که از تخم در میآیند در اتاقهای مولد نگهداری میکنند. خود باروری گاهی در گونه هایی از جنس Mnemiopsis دیده شده است [ 22 ] و تصور می شود که بیشتر گونه های هرمافرودیتی خود بارور هستند. [19]
رشد تخمهای بارور شده مستقیم است. هیچ شکل لارو مشخصی وجود ندارد. نوجوانان همه گروهها عموما پلانکتون هستند و بیشتر گونهها شبیه سیدیپیپیدهای بالغ مینیاتوری هستند و با رشد به تدریج فرمهای بدن بالغ خود را ایجاد میکنند. با این حال، در جنس Beroe ، جوجهها دهان بزرگ دارند و مانند بزرگسالان فاقد شاخک و غلاف شاخک هستند. در برخی از گروه ها، مانند پلاتیکتنیدهای مسطح و ساکن در پایین، جوجه ها بیشتر شبیه لاروهای واقعی رفتار می کنند. آنها در میان پلانکتونها زندگی میکنند و بنابراین جایگاه اکولوژیکی متفاوتی نسبت به والدین خود اشغال میکنند، و تنها پس از رها شدن به کف دریا، شکل بزرگسالی را با یک انتوژن رادیکالتر [22] به دست میآورند. [19]
حداقل در برخی از گونهها، کنتوفورهای جوان قادر به تولید مقادیر کمی تخمک و اسپرم هستند، در حالی که اندازه آنها بسیار کمتر است، و بالغین تا زمانی که غذای کافی دارند، تخمک و اسپرم تولید میکنند. اگر کمبود غذا داشته باشند، ابتدا تولید تخمک و اسپرم را متوقف می کنند و سپس اندازه آنها کوچک می شود. هنگامی که عرضه غذا بهبود می یابد، آنها به اندازه طبیعی رشد می کنند و سپس تولیدمثل را از سر می گیرند. این ویژگیها باعث میشود که کتنوفورها بتوانند جمعیت خود را خیلی سریع افزایش دهند. [19] اعضای Lobata و Cydippida نیز یک شکل تولید مثل دارند به نام dissogeny. دو مرحله بلوغ جنسی، ابتدا به عنوان لارو و بعداً به عنوان جوان و بالغ. آنها در طول مدتی که به عنوان لارو هستند قادرند گامت ها را به صورت دوره ای آزاد کنند. پس از پایان اولین دوره تولیدمثلی، آنها دیگر گامت های بیشتری تولید نخواهند کرد. جمعیتی از تخم مرتنزیا در دریای بالتیک مرکزی ، پدوژنتیک شده اند و فقط از لاروهای بالغ جنسی کمتر از 1.6 میلی متر تشکیل شده است. [66] [67]
در Mnemiopsis leidyi ، سیگنال دهی اکسید نیتریک (NO) هم در بافت های بالغ وجود دارد و هم به طور متفاوت در مراحل بعدی جنینی بیان می شود که نشان دهنده دخالت NO در مکانیسم های رشد است. [68] شکل بالغ همان گونه همچنین قادر است زمانی که توسط عوامل استرس زای محیطی تحریک می شود به مرحله سیدیپید بازگردد. [69]

اکثر کتنوفورهایی که در نزدیکی سطح زندگی می کنند، عمدتاً بی رنگ و تقریباً شفاف هستند. با این حال، برخی از گونههای عمیقتر دارای رنگدانههای قوی هستند، برای مثال گونهای که به عنوان «تورتوگاس قرمز» [70] شناخته میشود (به تصویر اینجا مراجعه کنید)، که هنوز به طور رسمی توضیح داده نشده است. [19] پلاتیکتنیدها عموماً متصل به سایر موجودات ته دریا زندگی می کنند و اغلب رنگ های مشابهی با این موجودات میزبان دارند. [19] روده جنس Bathocyroe در اعماق دریا قرمز است، که درخشندگی زیستی غلافهایی را که بلعیده است پنهان میکند. [56]
ردیفهای شانهای اکثر کتنوفورهای پلانکتونیک اثر رنگین کمانی ایجاد میکنند که ناشی از بیولومینسانس نیست، بلکه به دلیل پراکندگی نور در حین حرکت شانهها است. [19] [71] بیشتر گونهها نیز نورانی هستند، اما نور معمولاً آبی یا سبز است و فقط در تاریکی قابل مشاهده است. [19] با این حال، برخی از گروههای مهم، از جمله همه پلاتیکتنیدهای شناخته شده و جنس cydippid Pleurobrachia ، قادر به بیولومینسانس نیستند. [72]
هنگامی که برخی از گونهها، از جمله Bathyctena chuni ، Euplokamis stationis و Eurhamphaea vexilligera دچار اختلال میشوند، ترشحاتی (جوهر) تولید میکنند که تقریباً با طول موجهایی مشابه بدنشان درخشنده است. نوجوانان نسبت به بزرگسالان که درخشندگی آنها بر روی بدنشان پخش می شود، نسبت به اندازه بدنشان روشن تر می شوند. تحقیقات آماری دقیق عملکرد بیولومینسانس ctenophore ها را پیشنهاد نکرده است و هیچ ارتباطی بین رنگ دقیق آن و هر جنبه ای از محیط های حیوانات، مانند عمق یا اینکه آیا آنها در آب های ساحلی یا میانی اقیانوس زندگی می کنند، ایجاد نکرده است. [73]
در ctenophores، بیولومینسانس توسط فعال شدن پروتئین های فعال شده با کلسیم به نام فوتوپروتئین ها در سلول هایی به نام فوتوسیت ایجاد می شود ، که اغلب به کانال های نصف النهاری که زیر هشت ردیف شانه قرار دارند، محدود می شوند. در ژنوم Mnemiopsis leidyi ده ژن فتوپروتئین ها را کد می کنند. این ژنها با ژنهای اپسین در فتوسیتهای در حال رشد Mnemiopsis leidyi بیان میشوند و این احتمال را افزایش میدهد که تولید نور و تشخیص نور ممکن است با هم در این حیوانات کار کنند. [74]

Ctenophore ها در بیشتر محیط های دریایی یافت می شوند: از آب های قطبی در -2 درجه سانتی گراد تا مناطق استوایی در 30 درجه سانتی گراد. نزدیک سواحل و در وسط اقیانوس؛ از آب های سطحی تا اعماق اقیانوس در بیش از 7000 متر. [75] بهترینها جنسهای Pleurobrachia ، Beroe و Mnemiopsis هستند ، زیرا این فرمهای ساحلی پلانکتون یکی از محتملترین گونههایی هستند که در نزدیکی ساحل جمعآوری میشوند. [31] [56] هیچ گونه ctenophore در آب شیرین یافت نشده است.
در سال 2013 Mnemiopsis در دریاچه بیرکت قارون و در سال 2014 در دریاچه ال رایان II، هر دو در نزدیکی فیوم در مصر، که به طور تصادفی با حمل و نقل بچه ماهی (کفال) معرفی شدند، ثبت شد . اگرچه بسیاری از گونهها آبهای شور مانند مصبها و تالابهای ساحلی را در ارتباط باز با دریا ترجیح میدهند، این اولین رکورد از یک محیط داخلی بود. هر دو دریاچه شور هستند و Birket Qarun فوق شور است و نشان می دهد که برخی از ctenophore ها می توانند بدون اتصال به اقیانوس خود را در محیط های لیمنیک شور مستقر کنند. در دراز مدت انتظار نمی رود که جمعیت ها زنده بمانند. دو عامل محدود کننده در دریاچه های شور عبارتند از در دسترس بودن غذا و رژیم غذایی متنوع و درجه حرارت بالا در تابستان های گرم. از آنجایی که یک ایزوپود انگلی، Livoneca redmanii ، در همان زمان معرفی شد، دشوار است که بگوییم چقدر از تأثیرات اکولوژیکی گونههای مهاجم تنها توسط ctenophore ایجاد میشود. [76] [77]
Ctenophore ها ممکن است در طول ماه های تابستان در برخی از مناطق ساحلی فراوان باشند، اما در جاهای دیگر، غیر معمول و به سختی یافت می شوند.
در خلیجهایی که تعداد آنها بسیار زیاد است، شکار توسط ctenophores ممکن است جمعیت موجودات زئوپلانکتونیک کوچک مانند غلافهای کوچک را کنترل کند که در غیر این صورت ممکن است فیتوپلانکتونها (گیاهان پلانکتون) را که بخش حیاتی زنجیرههای غذایی دریایی هستند، از بین ببرند .
تقریباً همه کتنوفورها شکارچی هستند - هیچ گیاهخواری وجود ندارد و فقط یک جنس تا حدی انگلی وجود دارد . [56] اگر غذا فراوان باشد، می توانند 10 برابر وزن خود در روز غذا بخورند. [78] در حالی که Beroe عمدتاً سایر ctenophore ها را شکار می کند، سایر گونه های آب های سطحی به شکار زئوپلانکتون ها (حیوانات پلانکتونیک) می پردازند که در اندازه های میکروسکوپی، از جمله نرم تنان و لارو ماهی، تا سخت پوستان بالغ کوچک مانند غلاف ، دوپایان ، و حتی کریل متغیر است . اعضای جنس Haeckelia چتر دریایی را شکار میکنند و نماتوکیستهای طعمه (سلولهای نیشزن) را به جای کلوبلاست در شاخکهای خود قرار میدهند . [19] Ctenophore ها در طیف گسترده ای از تکنیک های خود برای گرفتن طعمه با عنکبوت ها مقایسه شده اند - برخی بدون حرکت در آب با استفاده از شاخک های خود به عنوان "تار" آویزان می شوند، برخی شکارچیان کمین مانند عنکبوت های جهنده Salticid هستند ، و برخی نیز قطره ای چسبنده را به سمت خود آویزان می کنند. انتهای یک نخ ظریف، همانطور که عنکبوت های بولاس انجام می دهند. این تنوع طیف گسترده ای از فرم های بدن را در یک شاخه با گونه های نسبتاً کمی توضیح می دهد. [56] Lampea "cydippid" دو شاخکدار منحصراً از سالپ ها تغذیه می کند ، خویشاوندان نزدیک سرپه های دریایی که مستعمرات شناور زنجیره ای بزرگی را تشکیل می دهند، و بچه های لامپیا مانند انگل خود را به نمک هایی می چسبانند که برای بلعیدن آنها خیلی بزرگ است. [56] اعضای جنس cydippid Pleurobrachia و Lobate Bolinopsis اغلب در یک مکان و زمان به تراکم جمعیت بالایی می رسند زیرا در انواع مختلف طعمه تخصص دارند: شاخک های بلند Pleurobrachia عمدتا شناگران نسبتاً قوی مانند غلاف های بالغ را می گیرند، در حالی که Bolinopsis به طور کلی از شناگران کوچکتر و ضعیفتر مانند روتیفرها و لاروهای نرم تنان و سخت پوستان تغذیه می کند . [79]
Ctenophores در گذشته به عنوان "بن بست" در زنجیره های غذایی دریایی در نظر گرفته می شد زیرا تصور می شد نسبت کم مواد آلی آنها به نمک و آب آنها را به رژیم غذایی نامناسبی برای حیوانات دیگر تبدیل می کند. همچنین اغلب اوقات شناسایی بقایای تنوفورها در روده شکارچیان احتمالی دشوار است، اگرچه شانه ها گاهی به اندازه کافی دست نخورده باقی می مانند تا سرنخی ارائه کنند. بررسی دقیق ماهی قزل آلا ، Oncorhynchus keta ، نشان داد که این ماهیها کانوفورها را 20 برابر سریعتر از وزنی برابر میگو هضم میکنند ، و اگر کتنوفورها به اندازه کافی در اطرافشان وجود داشته باشد، میتوانند رژیم غذایی خوبی ارائه دهند. بروئیدها عمدتاً در سایر کتنوفورها شکار می شوند. برخی از عروسهای دریایی و لاکپشتها مقادیر زیادی ctenophores میخورند و چتر دریایی ممکن است به طور موقت جمعیت ctenophore را از بین ببرد. از آنجایی که کتنوفورها و چتر دریایی ها اغلب دارای تنوع فصلی زیادی در جمعیت هستند، بیشتر ماهیانی که آنها را شکار می کنند عمومی هستند و ممکن است نسبت به ژله خواران متخصص تأثیر بیشتری بر روی جمعیت داشته باشند. این امر با مشاهده ماهیهای گیاهخوار که عمداً از زئوپلانکتونهای ژلاتینی در طول شکوفهدهی در دریای سرخ تغذیه میکنند، تأکید میکند. [80] لارو برخی از شقایق های دریایی انگل روی ctenophore ها هستند، همانطور که لارو برخی از کرم های مسطح که ماهی ها را هنگام رسیدن به بلوغ انگلی می کنند. [81]
بیشتر گونه ها هرمافرودیت هستند و بچه های جوان حداقل برخی از گونه ها قبل از رسیدن به اندازه و شکل بالغ قادر به تولید مثل هستند. این ترکیب هرمافرودیتیسم و تولید مثل اولیه جمعیت های کوچک را قادر می سازد با سرعت انفجاری رشد کنند.

Ctenophores ممکن است اکوسیستم های دریایی را با جلوگیری از فراوانی بیش از حد غلاف از خوردن تمام فیتوپلانکتون ها (گیاهان پلانکتون)، [82] که تولید کنندگان غالب مواد آلی دریایی از مواد غیر آلی هستند، متعادل کند. [83]
از سوی دیگر، در اواخر دهه 1980، ctenophore اقیانوس اطلس غربی Mnemiopsis leidyi به طور تصادفی از طریق مخازن بالاست کشتی ها به دریای سیاه و دریای آزوف وارد شد و به دلیل ایجاد افت شدید در صید ماهی با خوردن لارو و لارو ماهی مقصر شناخته شد. سخت پوستان کوچکی که در غیر این صورت ماهی بالغ را تغذیه می کنند. [82] Mnemiopsis به خوبی برای حمله به سرزمینهای جدید مجهز است (اگرچه این پیشبینی نشده بود تا زمانی که با موفقیت دریای سیاه را مستعمره کرد)، زیرا میتواند به سرعت رشد کند و طیف وسیعی از دما و شوری آب را تحمل کند . [84] این تأثیر توسط صید بیش از حد مزمن، و توسط اتروفیکاسیون که به کل اکوسیستم یک رونق کوتاه مدت داد، افزایش یافت و باعث شد که جمعیت Mnemiopsis حتی سریعتر از حد معمول افزایش یابد [85] - و مهمتر از همه به دلیل عدم وجود شکارچیان کارآمد در آنها. ctenophore ها را معرفی کرد. [84] جمعیت Mnemiopsis در آن مناطق در نهایت با معرفی تصادفی Ctenophore Beroe ovata آمریکای شمالی خوار Mnemiopsis ، [86] و با سرد شدن آب و هوای محلی از 1991 تا 1993، [85] که به طور قابل توجهی کند شد، تحت کنترل قرار گرفتند. متابولیسم حیوان [84] با این حال، بعید به نظر می رسد که فراوانی پلانکتون در این منطقه به سطوح قبل از Mnemiopsis بازگردانده شود . [87]
در اواخر دهه 1990 Mnemiopsis در دریای خزر ظاهر شد . Beroe ovata اندکی بعد وارد شد و انتظار می رود تاثیر Mnemiopsis را در آنجا کاهش دهد، اما نه از بین ببرد. Mnemiopsis همچنین در اواخر دهه 1990 به شرق مدیترانه رسید و اکنون به نظر می رسد در دریای شمال و دریای بالتیک در حال رشد است . [19]
تعداد گونههای ctenophore زنده نامشخص است، زیرا بسیاری از آنهایی که نامگذاری شده و به طور رسمی توصیف شدهاند با گونههایی که با نامهای علمی دیگر شناخته میشوند یکسان هستند. کلودیا میلز تخمین میزند که حدود 100 تا 150 گونه معتبر وجود دارد که تکراری نیستند، و حداقل 25 گونه دیگر، عمدتاً در اعماق دریا، بهعنوان متمایز شناخته شدهاند اما هنوز با جزئیات کافی برای پشتیبانی از یک توصیف و نامگذاری رسمی تجزیه و تحلیل نشدهاند. [70]
نویسندگان اولیه به دلیل شباهتهای مورفولوژیکی بین این دو گروه، ctenophores را با cnidarians در یک شاخه واحد به نام Coelenterata ترکیب کردند. مانند cnidarians، بدن ctenophores از یک توده ژله تشکیل شده است که یک لایه سلول در خارج و لایه دیگر حفره داخلی را پوشانده است. با این حال، در ctenophores، این لایه ها دو سلول عمیق هستند، در حالی که لایه های cnidarians تنها یک سلول واحد هستند. Ctenophores همچنین به دلیل تکیه بر جریان آب از طریق حفره بدن برای هضم و تنفس و همچنین در داشتن یک شبکه عصبی غیرمتمرکز به جای مغز شبیه cnidarians هستند. مطالعات ژنومی نشان داده است که نورونهای Ctenophora، که از بسیاری جهات با سایر نورونهای حیوانی متفاوت هستند، بهطور مستقل از سایر حیوانات تکامل یافتهاند، [88] و افزایش آگاهی از تفاوتهای بین ژلههای شانهای و دیگر coelentarata، اخیرا را متقاعد کرده است. نویسندگان این دو را به عنوان فیلاهای جداگانه طبقه بندی می کنند . موقعیت ctenophores در شجره تکاملی حیوانات مدتها مورد بحث بوده است و دیدگاه اکثریت در حال حاضر، بر اساس فیلوژنتیک مولکولی ، این است که cnidarians و biterian ها بیشتر از هرکدام به ctenophore ها مرتبط هستند.

طبقه بندی سنتی ctenophore ها را به دو دسته تقسیم می کند ، آنهایی که شاخک دارند ( Tentaculata ) و آنهایی که بدون ( Nuda ). نودا فقط شامل یک راسته ( Beroida ) و خانواده ( Beroidae ) و دو جنس ، Beroe (چند گونه) و Neis (یک گونه) است. [70]
Tentaculata به هشت راسته زیر تقسیم می شود : [ 70]
جنس های فسیلی وجود دارد که گروه ساقه ای در نظر گرفته می شود .
با وجود بدنهای شکننده و ژلاتینیشان، فسیلهایی که تصور میشود نمایانگر تنوفورها هستند - ظاهراً بدون شاخک اما ردیفهای شانهای بسیار بیشتر از اشکال مدرن - در لاگرشتن در اوایل دوران کامبرین ، حدود 515 میلیون سال پیش ، یافت شدهاند . با این وجود، تجزیه و تحلیل فیلوژنتیک مولکولی اخیر به این نتیجه می رسد که جد مشترک تقریباً 350 میلیون سال پیش 88 ± 88 میلیون سال پیش به وجود آمده است، که با تخمین های قبلی که نشان می دهد 66 میلیون سال پیش پس از رویداد انقراض کرتاسه-پالئوژن رخ داده است، در تضاد است . [89]
به دلیل بدن نرم و ژلاتینیشان، کتنوفورها بهعنوان فسیل بسیار کمیاب هستند و فسیلهایی که بهعنوان ctenophore تفسیر شدهاند، تنها در Lagerstätten یافت شدهاند ، مکانهایی که محیط بهطور استثنایی برای حفظ بافت نرم مناسب بود. تا اواسط دهه 1990، تنها دو نمونه به اندازه کافی برای تجزیه و تحلیل شناخته شده بود، هر دو عضو گروه تاج، از اوایل دوره دونین (Emsian) . سپس سه گونه فرضی دیگر در شیل برگس و سایر سنگهای کانادایی با سن مشابه، حدود 505 میلیون سال پیش در اواسط دوره کامبرین یافت شد . هر سه فاقد شاخک بودند اما بین 24 تا 80 ردیف شانه داشتند که بسیار بیشتر از هشت نوع معمولی گونه های زنده بود. همچنین به نظر می رسد که آنها ساختارهای اندام مانندی داشته اند که برخلاف آنچه در کتنوفورهای زنده یافت می شود. یکی از گونههای فسیلی که برای اولین بار در سال 1996 گزارش شد، دهان بزرگی داشت که ظاهراً توسط لبهای تاخورده احاطه شده بود که احتمالاً عضلانی بوده است. [3] شواهدی از چین یک سال بعد نشان می دهد که چنین ctenophore ها در کامبرین گسترده بودند، اما شاید بسیار متفاوت از گونه های امروزی بودند - برای مثال ردیف های شانه یک فسیل روی پره های برجسته نصب شده بود. [90] جوانترین فسیل یک گونه خارج از گروه تاج گونه Daihuoides متعلق به اواخر دونین است که متعلق به یک گروه پایه است که تصور می شد بیش از 140 میلیون سال قبل منقرض شده است. [91]
Ediacaran Eoandromeda می تواند ظاهراً نشان دهنده یک ژله شانه باشد. [4] دارای تقارن هشت برابری است، با هشت بازوی مارپیچی شبیه به ردیفهای شانهمانند یک تنوفور. اگر واقعاً ctenophore باشد، گروه را به مبدأ بیلاتریا نزدیک می کند. [92] فسیل اولیه کامبرین مانند ساقه بیپایان ، Stromatoveris ، متعلق به Chengjiang lagerstätte چین و قدمت آن به حدود 515 میلیون سال پیش ، بسیار شبیه به Vendobionta در دوره Ediacaran قبلی است . دی گان شو، سایمون کانوی موریس ، و همکاران. روی شاخههای آن، ردیفهایی از مژهها را یافتند که برای تغذیه فیلتر استفاده میشد . آنها پیشنهاد کردند که استروماتووریس یک "خاله" تکاملی از ctenophores است و ctenophore ها از حیوانات بی تحرک منشأ می گیرند که فرزندان آنها شناگر شده اند و مژک ها را از مکانیسم تغذیه به یک سیستم محرکه تغییر می دهند. [۹۳] فسیلهای دیگر کامبرین که از ایدهی اکتنوفورها که از شکلهای بیپایان تکامل یافتهاند حمایت میکنند، Dinomischus ، Daihua ، Xianguangia و Siphusauctum هستند که در کف دریا نیز زندگی میکردند، دارای اسکلتهای ارگانیک و شاخکهای پوشیده از مژگان در اطراف دهان خود بودند که توسط clad پیدا شده است. تجزیه و تحلیل به عنوان اعضای گروه ساقه ctenophore [94] [95]
فسیلهای کامبرین 520 میلیون ساله نیز از Chengjiang در چین نشان میدهند که یک کلاس کاملاً منقرض شده از ctenophore به نام " Scleroctenophora " که دارای یک اسکلت داخلی پیچیده با خارهای بلند بود. [96] این اسکلت همچنین از هشت فلپ با بدن نرم پشتیبانی می کرد که می توانست برای شنا و احتمالاً تغذیه استفاده شود. یک شکل، Thaumactena ، بدنی روان شبیه به کرمهای تیری داشت و میتوانست یک شناگر چابک باشد. [5]
رابطه فیلوژنتیکی ctenophores با بقیه Metazoa برای درک ما از تکامل اولیه حیوانات و منشا چند سلولی بسیار مهم است. سال هاست که کانون بحث بوده است. Ctenophores به عنوان دودمان خواهر Bilateria ، [ 97] [98] خواهر Cnidaria ، [99] [100] [101] [102] خواهر Cnidaria ، Placozoa ، و Bilateria ، [103] [104 ] ادعا شده است. ] [105] و خواهر با تمام حیوانات دیگر. [9] [106]
والتر گارستانگ در کتاب اشکال لاروی و دیگر آیات جانورشناسی خود ( Mülleria and the Ctenophore ) حتی نظریه ای را بیان کرد مبنی بر اینکه ctenophore ها از یک لارو Mülleria نئوتنیک چندکلاسی منشأ گرفته اند .
مجموعه ای از مطالعات که به حضور و غیاب اعضای خانواده های ژنی و مسیرهای سیگنالینگ (به عنوان مثال، هومئوباکس ها ، گیرنده های هسته ای ، مسیر سیگنال دهی Wnt و کانال های سدیم ) پرداختند، شواهدی مطابق با دو سناریوی اخیر نشان دادند که نشان می دهد کتنوفورها هر دو خواهر هستند. به Cnidaria ، Placozoa ، و Bilateria یا خواهر با تمام فیلاهای حیوانی دیگر. [107] [108] [109] [110] چندین مطالعه جدیدتر که ژنوم توالییابیشده کامل ctenophores را با دیگر ژنومهای حیوانی توالییابی شده مقایسه میکنند، همچنین از ctenophoreها بهعنوان دودمان خواهر همه حیوانات دیگر پشتیبانی میکنند. [111] [28] [112] [113] این موقعیت نشان میدهد که انواع سلولهای عصبی و ماهیچهای یا در دودمانهای اصلی جانوری از بین رفتهاند (مثلاً Porifera و Placozoa ) یا بهطور مستقل در دودمان ctenophore تکامل یافتهاند. [111]
محققان دیگر استدلال کرده اند که قرار گرفتن Ctenophora به عنوان خواهر برای تمام حیوانات دیگر یک ناهنجاری آماری ناشی از سرعت بالای تکامل در ژنوم ctenophore است و در عوض پوریفرا (اسفنج ها) اولین تاکسون حیوانی است که واگرا شده است. [105] [114] [115] [116] [117] آنها همچنین دارای نرخ بسیار بالایی از تکامل میتوکندری ، [118] و کمترین محتوای RNA/پروتئین شناخته شده ژنوم mtDNA در حیوانات هستند. [119] به این ترتیب، Ctenophora به نظر می رسد یک کلاد دیپلوبلاست پایه باشد . در توافق با نکته اخیر، تجزیه و تحلیل یک هم ترازی توالی بسیار بزرگ در مقیاس طبقه بندی متازوئن (1719 پروتئین در مجموع حدود 400000 موقعیت اسید آمینه ) نشان داد که کتنوفورها به عنوان دومین اصل و نسب حیوانات منشعب ظاهر می شوند، و اسفنج ها خواهر هستند. گروه به تمام حیوانات چند سلولی دیگر [8] همچنین، تحقیقات بر روی ژنهای موسین ، که به حیوان اجازه تولید مخاط را میدهد، نشان میدهد که اسفنجها هرگز آنها را نداشتهاند، در حالی که به نظر میرسد همه حیوانات دیگر، از جمله ژلههای شانه، ژنهایی با منشا مشترک دارند. [120] و مشخص شد که علیرغم تمام تفاوتهایشان، نورونهای ctenophoran همان پایه و اساس نورونهای cnidarian را دارند، پس از یافتهها نشان داد که نورونهای بیانکننده پپتید احتمالاً اجدادی برای انتقالدهندههای عصبی شیمیایی هستند. [121]
با این حال مطالعه دیگری به شدت این فرضیه را که اسفنج ها گروه خواهر همه حیوانات موجود هستند را رد می کند و قرار دادن Ctenophora را به عنوان گروه خواهر برای همه حیوانات دیگر ثابت می کند، و مخالفت با مقاله اخیر با مشکلات روش شناختی در تحلیل ها در آن توضیح داده می شود. کار کردن [13] نه کتنوفورها و نه اسفنجها دارای مسیرهای HIF نیستند ، [122] ژنوم آنها بر خلاف سایر حیوانات که دارای سه نوع هستند، تنها یک نوع کانال کلسیمی با ولتاژ را بیان میکنند، [123] و آنها تنها شاخههای حیوانی شناختهشدهای هستند که هیچ گونه درستی ندارند. ژن هاکس . [28] چند گونه از شاخه های دیگر. لارو nemertean pilidium، لارو گونه Phoronid Phoronopsis harmeri و لارو کرم بلوط Schizocardium californicum ، در رشد لاروی خود نیز به ژن های هوکس وابسته نیستند، اما در طول دگردیسی برای رسیدن به شکل بالغ خود به آنها نیاز دارند. [124] [125] [126] ژنهای اینکسین که پروتئینهای مورد استفاده برای ارتباط بین سلولی در حیوانات را کد میکنند، به نظر میرسد که به طور مستقل در ctenophores تکامل یافتهاند. [127]
از آنجایی که همه کتنوفورهای مدرن به جز بروئیدها دارای لاروهای سیدیپی مانند هستند، به طور گسترده تصور می شود که آخرین جد مشترک آنها نیز شبیه سیدیپیدها بوده و دارای بدنی تخم مرغی شکل و یک جفت شاخک جمع شونده است. تجزیه و تحلیل صرفا مورفولوژیکی ریچارد هاربیسون در سال 1985 به این نتیجه رسید که سیدیپیدها تکفیلتیک نیستند، به عبارت دیگر شامل همه و فقط نوادگان یک اجداد مشترک منفرد نیستند که خود سیدیپیدی بود. در عوض او دریافت که خانوادههای سیدیپیدیهای مختلف شبیهتر به اعضای دیگر راستههای ctenophore هستند تا سایر cydippids. او همچنین پیشنهاد کرد که آخرین جد مشترک کتنوفورهای مدرن یا سیدیپید مانند یا بروئید مانند بوده است. [129] تجزیه و تحلیل فیلوژنی مولکولی در سال 2001، با استفاده از 26 گونه، از جمله 4 گونه اخیراً کشف شده، تأیید کرد که cydippids تکفیلتیک نیستند و به این نتیجه رسیدند که آخرین جد مشترک ctenophores مدرن شبیه cydippid است. همچنین مشخص شد که تفاوتهای ژنتیکی بین این گونهها بسیار کوچک است - به قدری کوچک که روابط بین Lobata، Cestida و Thalassocalycida نامشخص باقی مانده است. این نشان می دهد که آخرین جد مشترک ctenophore های مدرن نسبتاً جدید بوده است و شاید از رویداد انقراض کرتاسه-پالئوژن در 65.5 میلیون سال پیش جان سالم به در برده در حالی که دودمان های دیگر از بین رفته اند. هنگامی که تجزیه و تحلیل گسترده شد تا نمایندگان سایر فیلاها را نیز شامل شود ، به این نتیجه رسید که cnidarians احتمالاً بیشتر از هر دو گروه با ctenophores مرتبط با دو طرفه هستند، اما این تشخیص نامشخص است. [128] یک کلاد شامل مرتنزیا ، کریستفان و یوپلوکامیس ممکن است دودمان خواهر همه کتنوفورهای دیگر باشد. [130] [13]
زمانهای واگرایی تخمین زدهشده از دادههای مولکولی نشان میدهد که تقریباً چند میلیون سال پیش (Mya) کلادهای اصلی تنوع داشتند: 350 Mya برای Cydippida نسبت به سایر Ctenophora، و 260 Mya برای Platyctenida نسبت به Beroida و Lobata. [13]
مطالعه روی ژله های ترکیبی نوع پیام رسان را نشان می دهد که احتمالاً در سیستم عصبی اجدادی کار می کند.- به استناد مقاله ژورنال:
مغز ctenophore نشان می دهد که اگر تکامل دوباره آغاز شود، هوش دوباره ظاهر می شود زیرا طبیعت خود را تکرار می کند.- بخشی از بررسی تحقیقات تا Moroz و همکاران. (2014)
{{cite report}}: CS1 maint: URL نامناسب ( پیوند )- شامل تصاویر