در باستان شناسی ، شکارچی-گردآورنده غربی ( WHG ، همچنین به عنوان شکارچی-گردآورنده اروپای غربی ، شکارچی-گردآورنده اروپای غربی یا خوشه اوبرکاسل نیز شناخته می شود ) ( حدود 15000 تا 5000 BP ) یک جزء اجدادی متمایز از اجداد اروپاییان مدرن است که نشان دهنده یک جمعیت اجدادی اروپاییان مدرن است. شکارچیان میان سنگی که در اروپای غربی ، جنوبی و مرکزی ، از جزایر بریتانیا در غرب تا کارپات ها در شرق، به دنبال عقب نشینی ورقه یخی آخرین حداکثر یخبندان پراکنده شدند . [2] ارتباط نزدیکی دارد و گاهی مترادف با مفهوم خوشه ویلابرونا است ، که نام آن از غار ریپاری ویلابرونا در ایتالیا گرفته شده است، که از پایانه پلیستوسن اروپا شناخته شده است، که عمدتاً اجدادی به جمعیت های بعدی WHG است.
WHGها نسبت به شکارچیان اروپایی قبلی رابطه ژنتیکی نزدیکتری با مردمان باستانی و مدرن در خاورمیانه و قفقاز دارند. روابط دقیق آنها با گروههای دیگر تا حدودی مبهم است، با منشأ خوشه ویلابرونا احتمالاً جایی در مجاورت بالکان . خوشه Villabruna تقریباً 19000 سال پیش به شبه جزیره ایتالیا و ایبری گسترش یافته بود، با خوشه WHG متعاقباً در پایان پلیستوسن در حدود 14-12000 سال پیش در سراسر اروپای غربی گسترش یافت و تا حد زیادی جایگزین Magdalenians که قبلاً بر منطقه تسلط داشتند. [3] مجدلنیاییها عمدتاً از گروههای کرومگنون اروپای غربی قبلی هستند که بیش از 30000 سال پیش، قبل از آخرین حداکثر یخبندان، به این منطقه رسیده بودند.
WHG ها یکی از گروه های ژنتیکی اصلی در دوره پس از یخبندان اوایل هولوسن اروپا، همراه با شکارچیان-گردآورنده شرقی (EHG) در اروپای شرقی بودند. مرز بین WHGها و EHGها تقریباً از دانوب پایینی ، به سمت شمال در امتداد جنگلهای غربی Dnieper به سمت دریای بالتیک غربی می رفت . [2] EHGها عمدتاً از مخلوطی از اجداد مرتبط با WHG و اوراسیا شمالی باستان (ANE) تشکیل شده بودند. [3] اسکاندیناوی توسط شکارچیان-گردآورنده اسکاندیناوی (SHGs) ساکن بود که مخلوطی بین WHG و EHG بودند. [4] در شبه جزیره ایبری، شکارچیان-گردآورنده اولیه هولوسن از ترکیبی از اجداد WHG و کرومگنون ماگدالن (GoyetQ2) تشکیل شده بودند. [5]
زمانی که جمعیت اصلی در سراسر اروپا بود، WHGها تا حد زیادی با گسترش متوالی کشاورزان اروپایی اولیه (EEFs) با منشأ آناتولی در اوایل دوران نوسنگی جایگزین شدند ، که عموماً به دلیل آمیختگی با گروه های WHG در طول گسترش اروپایی خود، مقدار کمی از اجداد WHG را داشتند. در میان جمعیت های امروزی، اصل و نسب WHG در میان جمعیت های منطقه بالتیک شرقی رایج است. [6]
شکارچیان-گردآورنده غربی (WHG) به عنوان یک جزء اجدادی متمایز شناخته می شوند که در اصل و نسب اکثر اروپاییان مدرن نقش دارند . [7] بیشتر اروپایی ها را می توان به عنوان مخلوطی از WHG، EEF، و WSH از استپ پونتیک-کاسپین مدل کرد . [8] WHGها همچنین به دیگر گروههای باستانی مانند کشاورزان اروپایی اولیه (EEF)، که عمدتاً از تبار آناتولی بودند، کمک کردند. [7] با گسترش دوران نوسنگی ، EEF بر استخر ژنی در بیشتر نقاط اروپا تسلط یافت، اگرچه نسب WHG در اروپای غربی از نوسنگی اولیه تا نوسنگی میانی تجدید حیات کرد. [9]
WHG ها نشان دهنده یک تغییر جمعیت عمده در اروپا در پایان عصر یخبندان است، احتمالاً گسترش جمعیت به اروپای قاره ای، از پناهگاه های اروپای جنوب شرقی یا غرب آسیا . [10] تصور می شود که اجداد آنها از اوراسیاهای شرقی در حدود 40000 قبل از میلاد و از اوراسیاهای شمالی باستان (ANE) قبل از 24000 BP (تاریخ تخمینی سن پسر مالتا ) جدا شدند. این تاریخ متعاقباً با یافتههای سایت شاخ کرگدن یانا به حدود 38 کیلیا، مدت کوتاهی پس از واگرایی دودمان غرب-اوراسیا و شرق-اوراسیا، به عقبتر نشان داده شد. [7] [11] والینی و همکاران. 2022 استدلال می کند که الگوهای پراکندگی و شکاف دودمان های اوراسیا غربی زودتر از c. 38000 سال پیش، با نمونههای قدیمیتر اروپایی دوران پارینه سنگی بالایی ، مانند نمونههای یافت شده در غارهای Zlaty Kun ، Peștera cu Oase و Bacho Kiro ، که ارتباطی با شکارچیان غربی نداشتند، اما به اوراسیاهای شرقی باستان نزدیکتر بودند یا به هر دو پایه. [12]
روابط خوشه WHG/Villabruna با دیگر گروههای انسانی پارینه سنگی در اروپا و غرب آسیا مبهم و مشمول تفاسیر متناقض است. [3] یک مطالعه در سال 2022 پیشنهاد کرد که جمعیت WHG/Villabruna از نظر ژنتیکی از شکارچیان در خاورمیانه و قفقاز در حدود 26000 سال پیش، در طول آخرین حداکثر یخبندان ، جدا شده است . [13] ژنومهای WHG در مقایسه با اروپاییهای دوران پارینه سنگی قبلی مانند گراوتیان ، تمایل بیشتری به جمعیتهای باستانی و مدرن خاورمیانه نشان میدهند . میل به جمعیت های خاورمیانه باستان در اروپا پس از آخرین حداکثر یخبندان افزایش یافت که با گسترش اجداد WHG (ویلابرونا یا اوبرکاسل) ارتباط داشت. همچنین شواهدی مبنی بر جریان ژنی دو طرفه بین WHG و جمعیت خاورمیانه در اوایل 15000 سال پیش وجود دارد. بقایای مرتبط با WHG عمدتاً متعلق به هاپلوگروه های کروموزوم Y انسانی I-M170 با فرکانس کمتر C-F3393 (مخصوصاً کلاد C-V20/C1a2 ) است که معمولاً در میان بقایای قدیمی اروپای پارینه سنگی مانند Kostenki-14 و سونگیر . هاپلوگروپ پدری C-V20 هنوز هم در مردانی که در اسپانیای مدرن زندگی میکنند یافت میشود که گواه حضور دیرینه این دودمان در اروپای غربی است. خوشه ویلابرونا همچنین حامل Y-haplogroup R1b (R1b-L754) است که از هاپلوگروه R * اوراسیا شمالی باستان گرفته شده است ، که نشان دهنده "پیوند اولیه بین اروپا و لبه غربی کمربند استپی اوراسیا" است. [14] کروموزوم های میتوکندریایی آنها عمدتاً به هاپلوگروه U5 تعلق داشت . [3] [15] یک مطالعه در سال 2023 پیشنهاد کرد که خوشه ویلابرونا از اختلاط تقریباً مساوی یک اصل و نسب اوراسیا غربی با اجداد اوراسیا غربی که نزدیک به فرد 35000 ساله BK1653 از غار باچو کیرو در بلغارستان است، پدید آمده است. این اصل و نسب مربوط به BK1653 نیز به طور قابل توجهی (~59٪) اجدادی به خوشه Věstonice مشخصه گروه های کرومگنون تولید کننده Gravettian شرقی است که ممکن است منعکس کننده اجداد مشترک در منطقه بالکان باشد. [3]
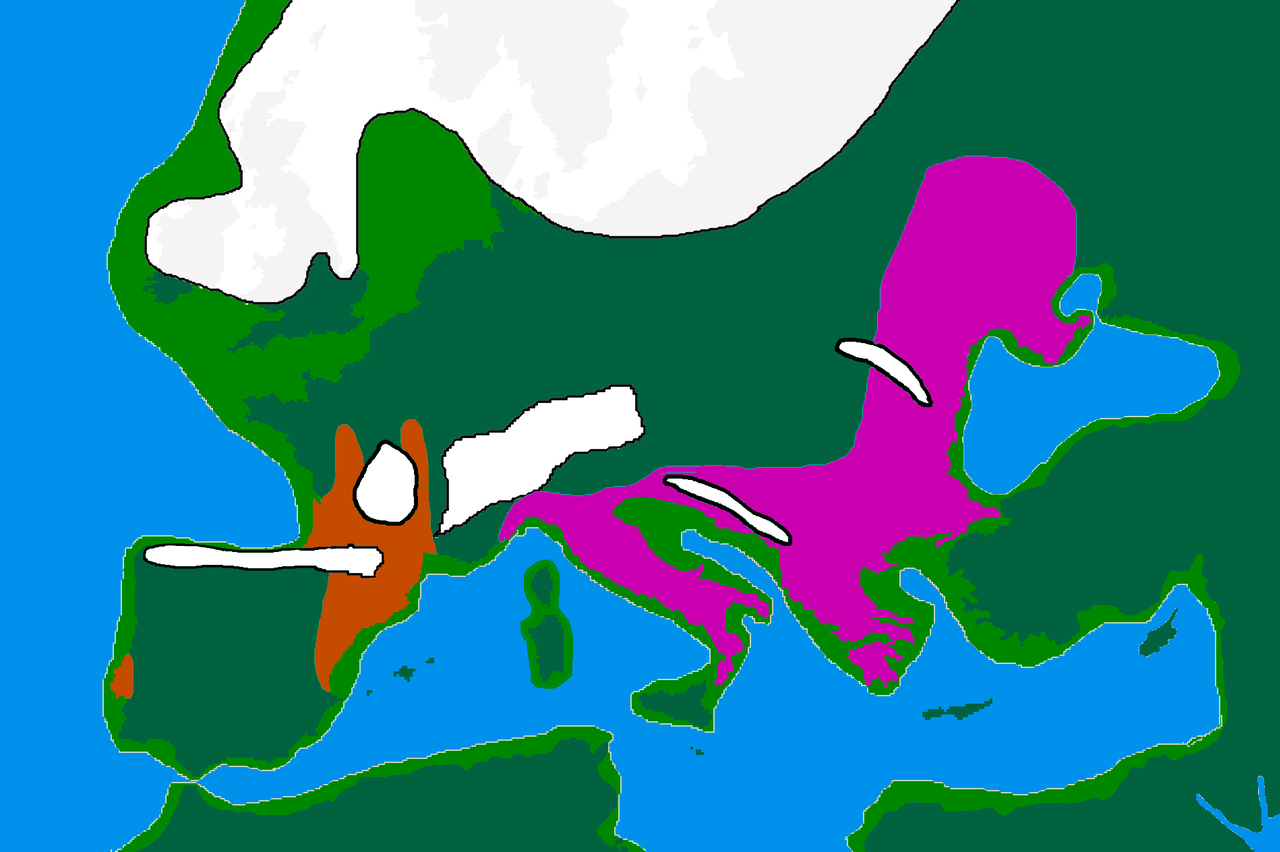
اولین افراد شناخته شده با اصل و نسب عمدتاً WHG/Villabruna در اروپا از ایتالیا شناخته شده اند و قدمت آنها به حدود 17000 سال پیش می رسد، [16] اگرچه فردی از غار El Mirón در شمال اسپانیا با 43٪ اجداد ویلابرونا مربوط به 19000 سال پیش است. در حالی که تایید نشده است، خوشه Villabruna احتمالا زودتر از منطقه بالکان نسبت به سایر نقاط اروپای جنوبی وجود داشته است. [3] جمعیتهای اولیه WHG/Villabruna با فرهنگ باستانشناسی Epigravettian مرتبط هستند ، که تا حد زیادی جایگزین جمعیتهای مرتبط با فرهنگ Magdalenian در حدود 14000 سال پیش شد (نسب خوشه Goyet-Q2 مرتبط با Magdalenian که عمدتاً از نسل Granttanve ، Soluttern قبلی است. گروه های تولید در فرانسه و اسپانیا). [3] [17] یک مطالعه در سال 2023 نشان داد که نسبت به جمعیتهای کرومگنون اروپای غربی قبلی مانند Gravettians، آن خوشه Goyet-Q2 مرتبط با Magdalenian دارای اصل و نسب چشمگیر (30٪) Villabruna حتی قبل از گسترش عمده WHG- بود. گروه های مرتبط در شمال آلپ این مطالعه همچنین نشان داد که نسبت به اعضای قبلی خوشه Villabruna از ایتالیا، گروههای مرتبط با WHG که در شمال کوههای آلپ از حدود 14000 سال پیش ظاهر شدند، حدود 25% از اجداد را از خوشه Goyet-Q2 (یا 10% از غرب) داشتند. خوشه فورنول مرتبط با گروتیان). این مقاله پیشنهاد می کند که WHG باید به نام خوشه Oberkassel ، به نام یکی از قدیمی ترین افراد WHG که در شمال کوه های آلپ یافت می شود، نامگذاری شود. [3] این مطالعه نشان میدهد که اصل و نسب Oberkassel عمدتاً قبل از گسترش، احتمالاً در اطراف غرب کوههای آلپ، به اروپای غربی و مرکزی و بریتانیا شکل گرفته است، جایی که افراد WHG نمونهبرداری شده از نظر ژنتیکی همگن هستند. این در تضاد با ورود اجداد ویلابرونا و اوبرکاسل به ایبریا است که به نظر میرسد درگیر رویدادهای مکرر اختلاط با جمعیتهای محلی با سطوح بالایی از نسب Goyet-Q2 است. این، و بقای کلادهای هاپلوگروپ C1 Y-DNA که قبلاً در میان شکارچی-گردآورندگان اولیه اروپایی مشاهده شده بود، نشان دهنده تداوم ژنتیکی نسبتاً بالاتر در جنوب غربی اروپا در این دوره است. [3]

همچنین مشخص شد که WHG به جمعیت هایی در مرزهای اروپا مانند کشاورزان اولیه آناتولی و آفریقایی های باستانی شمال غربی [19] و همچنین سایر گروه های اروپایی مانند شکارچیان شرقی کمک کرده است . [20] رابطه WHGها با EHGها قطعی نیست. [20] EHGها به گونهای مدلسازی میشوند که درجات مختلفی از اجداد را از دودمان مرتبط با WHG بدست آورند که از تنها 25٪ تا 91٪ متغیر است و بقیه به جریان ژنی از سیبریهای پارینه سنگی (ANE) و شاید شکارچی-جمعآوران قفقاز مرتبط است . یکی دیگر از دودمان معروف به شکارچی-گردآورندگان اسکاندیناوی (SHGs) ترکیبی از EHG و WHG است. [a] [22] [23]
در شبه جزیره ایبری، شکارچیان-گردآورنده هولوسن اولیه از جمعیت هایی با ترکیبی از اجداد WHG و کرومگنون ماگدالن (GoyetQ2) تشکیل شده بودند. [5]
مردم فرهنگ کوندای میان سنگی و فرهنگ ناروا در شرق بالتیک ترکیبی از WHG و EHG بودند، [24] که نزدیکترین قرابت را با WHG نشان میدهند. نمونههایی از دوران میانسنگی و نوسنگی اوکراین بهطور محکمی بین WHG و EHG جمع شدهاند که نشاندهنده تداوم ژنتیکی در رپیدز دنیپر برای یک دوره 4000 ساله است. نمونه های اوکراینی منحصراً متعلق به هاپلوگروپ مادری U بودند که در حدود 80 درصد از کل نمونه های شکارچی-گردآورنده اروپایی یافت می شود. [25]
مردم فرهنگ گودال شانه ای (CCC) در شرق بالتیک با EHG مرتبط بودند. [26] برخلاف بسیاری از WHGها، WHGهای شرق بالتیک در دوران نوسنگی ترکیب کشاورزان اروپایی را دریافت نکردند. بنابراین، جمعیتهای مدرن شرق بالتیک، نسبت به هر جمعیت دیگری در اروپا، دارای بیشتری از اجداد WHG هستند. [24]
مشخص شده است که SHGها حاوی ترکیبی از اجزای WHG هستند که احتمالاً از جنوب به اسکاندیناوی مهاجرت کرده اند و EHGهایی که بعداً از شمال شرقی در امتداد سواحل نروژ به اسکاندیناوی مهاجرت کرده اند . این فرضیه توسط شواهدی تأیید میشود که نشان میدهد SHGهای اسکاندیناوی غربی و شمالی نسبت به افراد اهل اسکاندیناوی شرقی (حدود 62%) اجداد WHG کمتری داشتند (حدود 51%). اعتقاد بر این است که WHGهایی که وارد اسکاندیناوی شده اند متعلق به فرهنگ Ahrensburg بوده اند . EHGها و WHGها فرکانس آللی کمتری SLC45A2 و SLC24A5 را نشان دادند که باعث ایجاد رنگدانه و OCA/Herc2 که باعث رنگ روشن چشم میشود ، نسبت به SHGها. [27]

DNA یازده WHG از پارینه سنگی فوقانی و میان سنگی در اروپای غربی ، اروپای مرکزی و بالکان ، با توجه به هاپلوگروههای Y-DNA و هاپلوگروههای mtDNA آنها مورد تجزیه و تحلیل قرار گرفت . تجزیه و تحلیل نشان داد که WHG ها زمانی به طور گسترده از سواحل اقیانوس اطلس در غرب، تا سیسیل در جنوب، تا بالکان در جنوب شرقی برای بیش از شش هزار سال توزیع شده بودند. [28] این مطالعه همچنین شامل تجزیه و تحلیل تعداد زیادی از افراد اروپای شرقی ماقبل تاریخ بود. سی و هفت نمونه از دوران میان سنگی و نوسنگی اوکراین (9500-6000 قبل از میلاد) جمع آوری شد. اینها به عنوان یک واسطه بین EHG و SHG تعیین شدند، اگرچه نسب WHG در این جمعیت در طول دوران نوسنگی افزایش یافت. نمونههای Y-DNA استخراجشده از این افراد منحصراً به هاپلوتیپهای R (بهویژه زیرشاخههای R1b1 ) و هاپلوتیپهای I (بهویژه زیرشاخههای I2 ) تعلق داشتند. mtDNA تقریباً به طور انحصاری به U (به ویژه زیرشاخه های U5 و U4 ) تعلق داشت. [28] تعداد زیادی از افراد از محل دفن Zvejnieki ، که بیشتر متعلق به فرهنگ کوندا و فرهنگ ناروا در شرق بالتیک بودند، مورد تجزیه و تحلیل قرار گرفتند. این افراد در مراحل اولیه عمدتاً از تبار WHG بودند، اما به مرور زمان اصل و نسب EHG غالب شد. Y-DNA این سایت تقریباً منحصراً به هاپلوتیپ های هاپلوگروپ R1b1a1a و I2a1 تعلق داشت . mtDNA منحصراً به هاپلوگروپ U (به ویژه زیرشاخه های U2 ، U4 و U5 ) تعلق داشت. [28] چهل فرد از سه مکان میان سنگی دروازههای آهنی در بالکان نیز مورد تجزیه و تحلیل قرار گرفتند. تخمین زده شد که این افراد دارای 85% WHG و 15% EHG هستند. نرها در این مکانها منحصراً هاپلوتیپهای هاپلوگروپ R1b1a و I (عمدتا زیرشاخههای I2a ) را حمل میکردند. mtDNA بیشتر به U (به ویژه زیرشاخه های U5 و U4 ) تعلق داشت. [28] مردم نوسنگی بالکان دارای 98٪ اجداد آناتولی و 2٪ اجداد WHG بودند. توسط کالکولیتیک ، مردم فرهنگ Cucuteni-Trypilliaمشخص شد که حدود 20 درصد اجداد شکارچی-گردآورنده را در خود جای داده است که حد وسط بین EHG و WHG بود. افراد فرهنگ آمفورای گلوبولار حدوداً در پناهگاه بودند. 25% اصل و نسب WHG، که به طور قابل توجهی بالاتر از گروه های نوسنگی میانی اروپای مرکزی است. [28]


یک مطالعه مهم در سال 2014 برای اولین بار سهم سه مولفه اصلی را در دودمان مدرن اروپایی شناسایی کرد: گردآورندگان شکارچی غربی (WHG، به نسبت 50٪ در اروپای شمالی)، اوراسیاهای شمالی باستان (ANE، سیبری های پارینه سنگی بالایی که بعداً با بعدها گسترش هند و اروپایی ، به نسبت 20٪، و در نهایت کشاورزان اروپایی اولیه (EEF، کشاورزانی از خاور نزدیک که از حدود 8000 BP به اروپا مهاجرت کردند، اکنون به نسبت 30٪ در بالتیک حضور دارند. منطقه به حدود 90٪ در مدیترانه). مولفه کشاورز اروپایی اولیه (EEF) بر اساس ژنوم یک زن دفن شده در سال شناسایی شد. 7000 سال پیش در یک گور فرهنگ خطی سفال در اشتوتگارت ، آلمان. [30]
این مطالعه در سال 2014 شواهدی مبنی بر اختلاط ژنتیکی بین WHG و EEF در سراسر اروپا پیدا کرد، با بیشترین سهم EEF در اروپای مدیترانه (به ویژه در ساردینیا، سیسیل، مالت و در میان یهودیان اشکنازی)، و بزرگترین سهم WHG در شمال اروپا و در میان باسک. مردم [31]
از سال 2014، مطالعات بیشتر تصویر آمیختگی بین EEF و WHG را اصلاح کرده است. در تجزیه و تحلیل 2017 از 180 مجموعه داده باستانی DNA مربوط به دوره های سنگی و نوسنگی از مجارستان، آلمان و اسپانیا، شواهدی از دوره طولانی آمیختگی پیدا شد. اختلاط به صورت منطقه ای، از جمعیت های شکارچی-گردآورنده محلی صورت گرفت، به طوری که جمعیت های سه منطقه (آلمان، ایبریا و مجارستان) از نظر ژنتیکی در تمام مراحل دوره نوسنگی قابل تشخیص بودند، با افزایش تدریجی نسبت اجداد WHG جمعیت های کشاورز در طول زمان. . این نشان می دهد که پس از گسترش اولیه کشاورزان اولیه، هیچ مهاجرت طولانی مدت دیگری به اندازه کافی برای یکنواخت کردن جمعیت کشاورز وجود نداشته است، و جمعیت کشاورز و شکارچی-گردآورنده قرن ها در کنار یکدیگر وجود داشته اند، با اختلاط تدریجی مداوم در طول قرن پنجم. تا هزاره چهارم قبل از میلاد (به جای یک رویداد ترکیبی در تماس اولیه). [32] نرخ های مخلوط از نظر جغرافیایی متفاوت بود. در اواخر نوسنگی، اجداد WHG در کشاورزان در مجارستان حدود 10٪، در آلمان حدود 25٪ و در ایبریا تا 50٪ بود. [33]
تجزیه و تحلیل بقایای از Grotta Continenza در ایتالیا نشان داد که از شش بقایای، سه بقایای آن بین سالهای قبل مدفون شده اند. 10000 سال قبل از میلاد و 7000 سال قبل از میلاد متعلق به I2a-P214 بود . و دو برابر هاپلوگروه های مادری U5b1 و یک U5b3 . [34] [35] حدود 6000 سال قبل از میلاد، WHGهای ایتالیا تقریباً به طور ژنتیکی با EEFs (دو G2a2 ) و یک هاپلوگروپ R1b جایگزین شدند ، اگرچه نسب WHG در هزارههای بعدی کمی افزایش یافت. [36]
افراد نوسنگی در جزایر بریتانیا نزدیک به جمعیت های دوران نوسنگی اولیه و میانی ایبری و اروپای مرکزی بودند، به گونه ای که حدود 75 درصد اجدادشان از EEF و بقیه از WHG در اروپای قاره ای مدل شده بودند. آنها متعاقباً اکثر جمعیت WHG را در جزایر بریتانیا بدون اختلاط زیاد با آنها جایگزین کردند. [37]
تخمین زده می شود که WHG بین 20 تا 30 درصد اجداد را به گروه های EEF نوسنگی در سراسر اروپا کمک کرده است. سازگاری های خاص در برابر پاتوژن های محلی ممکن است از طریق مخلوط WHG میانسنگی در جمعیت های EEF نوسنگی معرفی شده باشد. [38]
مطالعه ای بر روی شکارچیان میانسنگی از دانمارک نشان داد که آنها با شکارچیان-گردآورنده غربی معاصر مرتبط هستند و با فرهنگ های Maglemose ، Kongemose و Ertebølle مرتبط هستند . آنها "همگنی ژنتیکی را از حدود 10500 تا 5900 سال کالیبره شده قبل از امروز" نشان دادند تا اینکه "کشاورزان دوره نوسنگی با اجداد آناتولی وارد شدند". انتقال به دوره نوسنگی "بسیار ناگهانی بود و منجر به جابجایی جمعیت با مشارکت ژنتیکی محدود شکارچیان محلی شد. جمعیت بعدی نوسنگی با فرهنگ Funnelbeaker مرتبط شده است . [39]

به گفته دیوید رایش ، تجزیه و تحلیل DNA نشان داده است که گردآورندگان شکارچی غربی معمولاً پوست تیره، موهای تیره و چشم آبی داشتند. [42] پوست تیره به دلیل منشأ خارج از آفریقا بود (همه جمعیتهای هومو ساپینس در ابتدا پوست تیره داشتند)، در حالی که چشمهای آبی نتیجه تغییر در ژن OCA2 آنها بود که باعث بیرنگشدن عنبیه میشد . [43]
گریم وارن باستان شناس گفته است که رنگ پوست آنها از زیتونی تا سیاه متغیر است و حدس می زند که آنها ممکن است رنگ های چشم و موی منطقه ای داشته باشند. [44] این به طور قابل توجهی با شکارچیان-گردآورنده شرقی (EHG) که از دور مرتبط هستند متفاوت است - که گفته می شود پوست روشن، چشم قهوه ای یا چشم آبی و موهای تیره یا مو روشن هستند. [45]
دو اسکلت WHG با SNP ناقص ، La Braña و Cheddar Man ، پیش بینی می شود که دارای پوست تیره یا تیره تا سیاه باشند، در حالی که دو اسکلت WHG دیگر با SNP های کامل، "Sven" و Loschbour man ، پیش بینی می شود که دارای پوست تیره یا متوسط باشند. به ترتیب پوست تیره و میانی. [46] [27] [b] زیستشناس اسپانیایی کارلز لالوئزا-فاکس گفت که فرد La Braña-1 دارای پوست تیره است، "اگرچه ما نمیتوانیم سایه دقیق آن را بدانیم." [48]
طبق یک مطالعه در سال 2020، ورود کشاورزان اروپایی اولیه (EEFs) از غرب آناتولی از 8500 تا 5000 سال پیش، همراه با هردرهای استپ غربی در عصر برنز ، باعث تکامل سریع جمعیت های اروپایی به سمت پوست و موی روشن تر شد. [43] آمیختگی بین جمعیت شکارچی-گردآورنده و کشاورز ظاهراً گاه به گاه بود، اما گسترده نبود. [49]
.png/1280px-Archaeogenetic_analysis_of_human_skin_pigmentation_in_Europe_(with_Asia_geographic_extension).png)
برخی از نویسندگان در مورد بازسازی رنگدانه های پوست احتیاط کرده اند : Quillen و همکاران. (2019) مطالعاتی را تأیید می کند که به طور کلی نشان می دهد که "رنگ پوست روشن تر در بسیاری از اروپا در دوران میان سنگی غیرمعمول بود"، از جمله مطالعات در مورد پیش بینی های "تیره یا تیره تا سیاه" برای مرد چدار، اما هشدار می دهد که "بازسازی های میان سنگی و نوسنگی فنوتیپ رنگدانهای با استفاده از جایگاههای رایج در جمعیتهای مدرن باید با احتیاط تفسیر شود، زیرا ممکن است جایگاههای دیگری که هنوز بررسی نشدهاند نیز بر فنوتیپ تأثیر گذاشته باشند. [50]
سوزان والش، متخصص ژنتیک در دانشگاه ایندیانا – دانشگاه پردو ایندیاناپولیس ، که بر روی پروژه مرد چدار کار می کرد، گفت که ما رنگ پوست او را نمی دانیم. [51] بیوشیمیدان آلمانی یوهانس کراوز اظهار داشت که ما نمی دانیم که آیا رنگ پوست شکارچیان اروپای غربی بیشتر شبیه رنگ پوست مردم آفریقای مرکزی امروزی است یا مردم منطقه عرب . تنها مشخص است که آنها حامل هیچ جهش شناخته شده ای نیستند که مسئول پوست روشن در جمعیت های بعدی اروپایی ها باشد. [52]
یک تحقیق در سال 2024 در مورد اصل و نسب ژنومی و پویایی اجتماعی آخرین شکارچیان-گردآورنده اقیانوس اطلس فرانسه بیان کرده است که "از لحاظ فنوتیپی، ما در طول دوره میان سنگی پسین در فرانسه تنوعی پیدا می کنیم" که در آن دو مورد از WHG در این مطالعه "احتمالاً دارای توالی بودند. رنگدانههای پوست کمرنگ تا متوسط»، اما «اکثر افراد دارای پوست تیره و چشمهای آبی مشخصه WHG هستند» نمونههای مورد مطالعه. [53]
{{cite journal}}: CS1 maint: چندین نام: فهرست نویسندگان ( پیوند )با شروع خوشه ویلابرونا حداقل 14000 سال پیش، همه افراد اروپایی که مورد تجزیه و تحلیل قرار گرفتند، تمایلی به خاور نزدیک نشان دادند. این از نظر زمانی با بیناستادی Bølling-Allerød، اولین دوره گرم شدن قابل توجه پس از عصر یخبندان، مرتبط است. از نظر باستان شناسی، با انتقال فرهنگی در Epigravettian در جنوب اروپا و انتقال Magdalenian به Azilian در اروپای غربی ارتباط دارد. بنابراین، ظاهر خوشه ویلابرونا ممکن است منعکس کننده مهاجرت یا جابجایی جمعیت در اروپا در پایان عصر یخبندان باشد، مشاهداتی که با شواهد گردش توالی DNA میتوکندریایی در این زمان نیز مطابقت دارد. یکی از سناریوهایی که می تواند این الگوها را توضیح دهد، گسترش جمعیت از پناهگاه های جنوب شرق اروپا یا غرب آسیا پس از عصر یخبندان است که تبار ژنتیکی اروپا و خاور نزدیک را در کنار هم قرار می دهد. ششم، در خوشه ویلابرونا، برخی، اما نه همه، افراد به آسیای شرقی میل دارند. یک جهت مهم برای کار آینده، تولید داده های مشابه DNA باستانی از اروپای جنوب شرقی و خاور نزدیک است تا به تصویر کامل تری از تاریخ جمعیت پارینه سنگی بالایی اوراسیا غربی برسیم.
{{cite web}}: CS1 maint: نام های عددی: فهرست نویسندگان ( پیوند )از طریق این تجزیه و تحلیل، ما یک کروموزوم از جنوب اروپا را به عنوان یک شاخه عمیق جدید در هاپلوگروه C شناسایی کردیم (C-V20 یا C7، شکل S1). پیش از این، تنها چند نمونه از کروموزوم های C (فقط با نشانگر RPS4Y711 تعریف شده بود) در جنوب اروپا یافت شده بود [32]، [33]. برای بهبود دانش خود در مورد توزیع هاپلوگروه C در اروپا، 1965 فرد اروپایی را برای جهش RPS4Y711 بررسی کردیم و یک کروموزوم هاپلوگروپ C اضافی را از جنوب اروپا شناسایی کردیم که به عنوان C7 نیز طبقه بندی شده است (داده ها نشان داده نشده است).
بیشتر اروپاییهای امروزی حداقل از سه جمعیت بسیار متمایز سرچشمه میگیرند: شکارچیان اروپای غربی، که نسب خود را به همه اروپاییها رساندهاند، اما نه خاور نزدیک. اوراسیاهای شمالی باستان مربوط به سیبریان پارینه سنگی بالایی بودند که هم به اروپاییان و هم به خاور نزدیک کمک کردند. و کشاورزان اروپایی اولیه، که عمدتاً منشأ خاور نزدیک داشتند، اما همچنین دارای اجداد مربوط به شکارچی-گردآورنده اروپای غربی بودند.
انسان خردمند حدود 42000 سال پیش از خاور نزدیک وارد اروپا شد. این انسان ها مانند منشاء آفریقایی خود پوست تیره داشتند اما به دلیل تنوع ژن OCA2 (که باعث بی رنگ شدن عنبیه می شود) بسیاری از آنها چشمان آبی داشتند. 8500 تا 5000 سال پیش توسط کشاورزان غرب آناتولی معرفی شد. این شروع انقلاب نوسنگی در این مناطق بود که با سبک زندگی بی تحرک تر و اهلی کردن برخی گونه های جانوری و گیاهی مشخص می شد. (...) "افزایش سریع جمعیت به دلیل انقلاب نوسنگی، مانند استفاده از محصولات شیر به عنوان منبع غذایی برای بزرگسالان و ظهور کشاورزی، و همچنین گسترش گسترده دامداران یامنایا احتمالا باعث انتخاب سریع در جمعیت های اروپایی به سمت پوست و موهای روشن حرکت کنید
هنگامی که در آناتولی تأسیس شد، تیم Excoffier دریافتند، جمعیت کشاورزی اولیه به روشی شبیه به سنگ پله به سمت غرب به سمت اروپا نقل مکان کردند که از حدود 8000 سال پیش شروع شد. آنها گاهی - اما نه به طور گسترده - با شکارچیان محلی مخلوط می شدند.
تجزیه و تحلیلهای آنها نشان میدهد که رنگ پوست هر دو فرد احتمالاً تیره بوده است، و پیشبینی میشود که رنگ پوست چدار بینسنگی «تیره یا تیره تا سیاه» باشد. این یافتهها نشان میدهد که رنگ پوست روشنتر در بیشتر اروپا در دوران میان سنگی غیرمعمول بوده است. با این حال، این در تضاد با تخمین تاریخ کمتر از 20 کیا در بالا نیست، که به شروع انتخاب و نه زمان تثبیت آلل های مورد علاقه می پردازد (Beleza و همکاران، 2013؛ بلزا، جانسون، و همکاران، 2013). . در حالی که مطالعات ژنوم باستانی رنگ پوست عموماً تیرهتر را در میان اروپاییان میانسنگی پیشبینی میکردند، آللهای مشتقشده در rs1426654 و rs16891982 در جمعیتهای اروپایی در طول دوره میان سنگی جدا میشدند (González-Fortes et al., 2017; Günther et al., Mittniket, 2018; ، نشان می دهد که تنوع فنوتیپی ناشی از این مکان ها احتمالاً در این زمان وجود داشته است. با این حال، بازسازی فنوتیپ رنگدانهای میان سنگی و نوسنگی با استفاده از جایگاههای رایج در جمعیتهای مدرن باید با احتیاط تفسیر شود، زیرا ممکن است مکانهای دیگر هنوز بررسی نشده نیز بر فنوتیپ تأثیر گذاشته باشند.
_to_Villabruna_(orange)_genetic_types_in_European_sites,_according_to_timeline_and_climate_evolution.jpg/1280px-Transition_from_Goyet_(green)_to_Villabruna_(orange)_genetic_types_in_European_sites,_according_to_timeline_and_climate_evolution.jpg){kind=link}